
Abstract
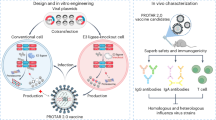
Generating effective live vaccines from intact viruses remains challenging owing to considerations of safety and immunogenicity. Approaches that can be applied in a systematic manner are needed. Here we created a library of live attenuated influenza vaccines by using diverse cellular E3 ubiquitin ligases to generate proteolysis-targeting (PROTAR) influenza A viruses. PROTAR viruses were engineered to be attenuated by the ubiquitin–proteasome system, which mediates viral protein degradation in conventional host cells, but allows efficient replication in engineered cell lines for large-scale manufacturing. Depending on the degron–E3 ligase pairs, viruses showed varying degrees of attenuation. In animal models, PROTAR viruses were highly attenuated and elicited robust, broad, strain-dependent humoral, mucosal and cellular immunity. In addition, they provided cross-reactive protection against homologous and heterologous viral challenges. This study provides a systematic approach for developing safe and effective vaccines, with potential applications in designing live attenuated vaccines against other pathogens.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

Data availability
All data associated with this study are available in the main text or Supplementary Information. The gene sequences of the WSN influenza virus strain used in this work have been deposited in GenBank under accession numbers CY034139.1, CY034138.1, X17336.1, HE802059.1, CY034135.1, CY034134.1, L25818.1 and CY034136.1. Data showing the expression of E3 ubiquitin ligases in lung tissues of humans and mice are available in Expression Atlas (https://www.ebi.ac.uk/gxa/home): KLHDC2 (https://www.ebi.ac.uk/gxa/genes/ensg00000165516?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000020978?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), KLHDC3 (https://www.ebi.ac.uk/gxa/genes/ensg00000124702?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000063576?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), APPBP2 (https://www.ebi.ac.uk/gxa/genes/ensg00000062725?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000018481?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), KLHL20 (https://www.ebi.ac.uk/gxa/genes/ensg00000076321?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000026705?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), FBXO31 (https://www.ebi.ac.uk/gxa/genes/ensg00000103264?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000052934?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), β-TrCP (https://www.ebi.ac.uk/gxa/genes/ensg00000166167?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000025217?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), SOCS2 (https://www.ebi.ac.uk/gxa/genes/ensg00000120833?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000020027?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), FEM1C (https://www.ebi.ac.uk/gxa/genes/ensg00000145780?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000033319?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), ITCH (https://www.ebi.ac.uk/gxa/genes/ensg00000078747?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000027598?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), SPOP (https://www.ebi.ac.uk/gxa/genes/ensg00000121067?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000057522?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), MDM2 (https://www.ebi.ac.uk/gxa/genes/ensg00000135679?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000020184?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), FEM1B (https://www.ebi.ac.uk/gxa/genes/ensg00000169018?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000032244?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D#baseline), CBL (https://www.ebi.ac.uk/gxa/genes/ensg00000110395?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline; https://www.ebi.ac.uk/gxa/genes/ensmusg00000034342?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), COP1 (https://www.ebi.ac.uk/gxa/genes/ensg00000143207?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D#baseline, https://www.ebi.ac.uk/gxa/genes/ensmusg00000040782?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), FBW7 (https://www.ebi.ac.uk/gxa/genes/ensg00000109670?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline, https://www.ebi.ac.uk/gxa/genes/ensmusg00000028086?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), Skp2 (https://www.ebi.ac.uk/gxa/genes/ensg00000145604?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D#baseline, https://www.ebi.ac.uk/gxa/genes/ensmusg00000054115?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), SIAH-1 (https://www.ebi.ac.uk/gxa/genes/ensg00000196470?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), KEAP1 (https://www.ebi.ac.uk/gxa/genes/ensg00000079999?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline, https://www.ebi.ac.uk/gxa/genes/ensmusg00000003308?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), APC/C (CDC20) (https://www.ebi.ac.uk/gxa/genes/ensg00000117399?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline, https://www.ebi.ac.uk/gxa/genes/ensmusg00000006398?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline), UBR4 (https://www.ebi.ac.uk/gxa/genes/ensg00000127481?bs=%7B%22homo%20sapiens%22%3A%5B%22ORGANISM_PART%22%5D%7D#baseline, https://www.ebi.ac.uk/gxa/genes/ensmusg00000066036?bs=%7B%22mus%20musculus%22%3A%5B%22ORGANISM_PART%22%5D%7D&ds=%7B%22kingdom%22%3A%5B%22animals%22%5D%7D#baseline). Data showing the expression of β-TrCP in human tissues are available in the Human Protein Atlas (https://www.proteinatlas.org/ENSG00000166167-BTRC/tissue). Data showing the expression of β-TrCP in diverse types of cells from the human respiratory system are available in the Human Protein Atlas (https://www.proteinatlas.org/ENSG00000166167-BTRC/single+cell+type). The RNA-seq data, scRNA-seq data and scBCR-seq data have been deposited in the CNGB Nucleotide Sequence Archive (accession code: CNP0005500) and the NCBI Sequence Read Archive (accession number PRJNA1193130). There is no restriction on data availability. Source data are provided with this paper.
Change history
12 June 2025
A Correction to this paper has been published: https://doi.org/10.1038/s41564-025-02053-0
References
Yamayoshi, S. & Kawaoka, Y. Current and future influenza vaccines. Nat. Med. 25, 212–220 (2019).
Iuliano, A. D. et al. Estimates of global seasonal influenza-associated respiratory mortality: a modelling study. Lancet 391, 1285–1300 (2018).
Wei, C. J. et al. Next-generation influenza vaccines: opportunities and challenges. Nat. Rev. Drug Discov. 19, 239–252 (2020).
Gouma, S., Anderson, E. M. & Hensley, S. E. Challenges of making effective influenza vaccines. Annu Rev. Virol. 7, 495–512 (2020).
Jang, Y. H. & Seong, B. L. Principles underlying rational design of live attenuated influenza vaccines. Clin. Exp. Vaccin. Res. 1, 35–49 (2012).
Blanco-Lobo, P., Nogales, A., Rodriguez, L. & Martinez-Sobrido, L. Novel approaches for the development of live attenuated influenza vaccines. Viruses 11, 190 (2019).
Mueller, S. et al. Live attenuated influenza virus vaccines by computer-aided rational design. Nat. Biotechnol. 28, 723–726 (2010).
Stech, J., Garn, H., Wegmann, M., Wagner, R. & Klenk, H. D. A new approach to an influenza live vaccine: modification of the cleavage site of hemagglutinin. Nat. Med. 11, 683–689 (2005).
Bournazos, S. & Ravetch, J. V. Attenuated vaccines for augmented immunity. Cell Host Microbe 21, 314–315 (2017).
Moeller, A., Kirchdoerfer, R. N., Potter, C. S., Carragher, B. & Wilson, I. A. Organization of the influenza virus replication machinery. Science 338, 1631–1634 (2012).
Bekes, M., Langley, D. R. & Crews, C. M. PROTAC targeted protein degraders: the past is prologue. Nat. Rev. Drug Discov. 21, 181–200 (2022).
Tsai, J. M., Nowak, R. P., Ebert, B. L. & Fischer, E. S. Targeted protein degradation: from mechanisms to clinic. Nat. Rev. Mol. Cell Biol. 25, 740–757 (2024).
Timms, R. T. & Koren, I. Tying up loose ends: the N-degron and C-degron pathways of protein degradation. Biochem. Soc. Trans. 48, 1557–1567 (2020).
Si, L. et al. Generation of a live attenuated influenza A vaccine by proteolysis targeting. Nat. Biotechnol. 40, 1370–1377 (2022).
Koren, I. et al. The eukaryotic proteome is shaped by E3 ubiquitin ligases targeting C-terminal degrons. Cell 173, 1622–1635.e14 (2018).
Rusnac, D. V. et al. Recognition of the diglycine C-end degron by CRL2KLHDC2 ubiquitin ligase. Mol. Cell 72, 813–822.e4 (2018).
Lin, H. C. et al. C-terminal end-directed protein elimination by CRL2 ubiquitin ligases. Mol. Cell 70, 602–613.e3 (2018).
Chen, Z., Picaud, S., Filippakopoulos, P., D’Angiolella, V. & Bullock, A. N. Structural basis for recruitment of DAPK1 to the KLHL20 E3 ligase. Structure 27, 1395–1404.e4 (2019).
Li, Y. et al. Structural basis of the phosphorylation-independent recognition of cyclin D1 by the SCFFBXO31 ubiquitin ligase. Proc. Natl Acad. Sci. USA 115, 319–324 (2018).
Wu, G. et al. Structure of a β-TrCP1–Skp1–β-catenin complex: destruction motif binding and lysine specificity of the SCFβ-TrCP1 ubiquitin ligase. Mol. Cell 11, 1445–1456 (2003).
Kung, W. W., Ramachandran, S., Makukhin, N., Bruno, E. & Ciulli, A. Structural insights into substrate recognition by the SOCS2 E3 ubiquitin ligase. Nat. Commun. 10, 2534 (2019).
Liu, Y. et al. Structural basis for the regulatory role of the PPxY motifs in the thioredoxin-interacting protein TXNIP. Biochem. J. 473, 179–187 (2016).
Zhuang, M. et al. Structures of SPOP–substrate complexes: insights into molecular architectures of BTB-Cul3 ubiquitin ligases. Mol. Cell 36, 39–50 (2009).
Nicholson, J. et al. A systems wide mass spectrometric based linear motif screen to identify dominant in-vivo interacting proteins for the ubiquitin ligase MDM2. Cell Signal. 26, 1243–1257 (2014).
Kussie, P. H. et al. Structure of the MDM2 oncoprotein bound to the p53 tumor suppressor transactivation domain. Science 274, 948–953 (1996).
Chen, X. et al. Molecular basis for arginine C-terminal degron recognition by Cul2FEM1 E3 ligase. Nat. Chem. Biol. 17, 254–262 (2021).
Di Fiore, B. et al. The ABBA motif binds APC/C activators and is shared by APC/C substrates and regulators. Dev. Cell 32, 358–372 (2015).
Kapanidou, M., Lee, S. & Bolanos-Garcia, V. M. BubR1 kinase: protection against aneuploidy and premature aging. Trends Mol. Med. 21, 364–372 (2015).
Diaz-Martinez, L. A. et al. The Cdc20-binding Phe box of the spindle checkpoint protein BubR1 maintains the mitotic checkpoint complex during mitosis. J. Biol. Chem. 290, 2431–2443 (2015).
Ng, C. et al. Structural basis for a novel intrapeptidyl H-bond and reverse binding of c-Cbl-TKB domain substrates. EMBO J. 27, 804–816 (2008).
Hu, J. & Hubbard, S. R. Structural characterization of a novel Cbl phosphotyrosine recognition motif in the APS family of adapter proteins. J. Biol. Chem. 280, 18943–18949 (2005).
He, J. et al. Insights into degron recognition by APC/C coactivators from the structure of an Acm1–Cdh1 complex. Mol. Cell 50, 649–660 (2013).
Zur, A. & Brandeis, M. Securin degradation is mediated by fzy and fzr, and is required for complete chromatid separation but not for cytokinesis. EMBO J. 20, 792–801 (2001).
Pfleger, C. M. & Kirschner, M. W. The KEN box: an APC recognition signal distinct from the D box targeted by Cdh1. Genes Dev. 14, 655–665 (2000).
Karamysheva, Z., Diaz-Martinez, L. A., Crow, S. E., Li, B. & Yu, H. Multiple anaphase-promoting complex/cyclosome degrons mediate the degradation of human Sgo1. J. Biol. Chem. 284, 1772–1780 (2009).
Leismann, O. & Lehner, C. F. Drosophila securin destruction involves a D-box and a KEN-box and promotes anaphase in parallel with Cyclin A degradation. J. Cell Sci. 116, 2453–2460 (2003).
Tian, W. et al. Structural analysis of human Cdc20 supports multisite degron recognition by APC/C. Proc. Natl Acad. Sci. USA 109, 18419–18424 (2012).
Uljon, S. et al. Structural basis for substrate selectivity of the E3 ligase COP1. Structure 24, 687–696 (2016).
Kung, J. E. & Jura, N. The pseudokinase TRIB1 toggles an intramolecular switch to regulate COP1 nuclear export. EMBO J. 38, e99708 (2019).
Davydov, I. V. & Varshavsky, A. RGS4 is arginylated and degraded by the N-end rule pathway in vitro. J. Biol. Chem. 275, 22931–22941 (2000).
Hu, R. G. et al. The N-end rule pathway as a nitric oxide sensor controlling the levels of multiple regulators. Nature 437, 981–986 (2005).
Varshavsky, A. The N-end rule pathway and regulation by proteolysis. Protein Sci. 20, 1298–1345 (2011).
Hao, B., Oehlmann, S., Sowa, M. E., Harper, J. W. & Pavletich, N. P. Structure of a Fbw7–Skp1–cyclin E complex: multisite-phosphorylated substrate recognition by SCF ubiquitin ligases. Mol. Cell 26, 131–143 (2007).
Welcker, M. et al. The Fbw7 tumor suppressor regulates glycogen synthase kinase 3 phosphorylation-dependent c-Myc protein degradation. Proc. Natl Acad. Sci. USA 101, 9085–9090 (2004).
Yada, M. et al. Phosphorylation-dependent degradation of c-Myc is mediated by the F-box protein Fbw7. EMBO J. 23, 2116–2125 (2004).
Hao, B. et al. Structural basis of the Cks1-dependent recognition of p27Kip1 by the SCFSkp2 ubiquitin ligase. Mol. Cell 20, 9–19 (2005).
House, C. M. et al. A binding motif for Siah ubiquitin ligase. Proc. Natl Acad. Sci. USA 100, 3101–3106 (2003).
Tiedt, R., Bartholdy, B. A., Matthias, G., Newell, J. W. & Matthias, P. The RING finger protein Siah-1 regulates the level of the transcriptional coactivator OBF-1. EMBO J. 20, 4143–4152 (2001).
Velasco, K. et al. An N-terminal SIAH-interacting motif regulates the stability of the ubiquitin specific protease (USP)-19. Biochem. Biophys. Res. Commun. 433, 390–395 (2013).
Lu, M. et al. Discovery of a Keap1-dependent peptide PROTAC to knockdown Tau by ubiquitination–proteasome degradation pathway. Eur. J. Med. Chem. 146, 251–259 (2018).
Glotzer, M., Murray, A. W. & Kirschner, M. W. Cyclin is degraded by the ubiquitin pathway. Nature 349, 132–138 (1991).
King, R. W. & Kirschner, G. M. MW mutagenic analysis of the destruction signal of mitotic cyclins and structural characterization of ubiquitinated intermediates. Mol. Biol. Cell 7, 1343–1357 (1996).
Barford, D. Structure, function and mechanism of the anaphase promoting complex (APC/C). Q. Rev. Biophys. 44, 153–190 (2011).
Rotin, D. & Kumar, S. Physiological functions of the HECT family of ubiquitin ligases. Nat. Rev. Mol. Cell Biol. 10, 398–409 (2009).
Si, L. et al. A human-airway-on-a-chip for the rapid identification of candidate antiviral therapeutics and prophylactics. Nat. Biomed. Eng. 5, 815–829 (2021).
Bai, H. et al. Mechanical control of innate immune responses against viral infection revealed in a human lung alveolus chip. Nat. Commun. 13, 1928 (2022).
Si, L. et al. Clinically relevant influenza virus evolution reconstituted in a human lung airway-on-a-chip. Microbiol. Spectr. 9, e0025721 (2021).
Si, L. et al. Self-assembling short immunostimulatory duplex RNAs with broad-spectrum antiviral activity. Mol. Ther. Nucleic Acids 29, 923–940 (2022).
Benam, K. H. et al. Small airway-on-a-chip enables analysis of human lung inflammation and drug responses in vitro. Nat. Methods 13, 151–157 (2016).
Yaghi, A. & Dolovich, M. B. Airway epithelial cell cilia and obstructive lung disease. Cells 5, 40 (2016).
Sun, X. et al. A census of the lung: CellCards from LungMAP. Dev. Cell 57, 112–145.e2 (2022).
Hahn, S., Setz, C., Wild, J. & Schubert, U. The PTAP sequence within the p6 domain of human immunodeficiency virus type 1 Gag regulates its ubiquitination and MHC class I antigen presentation. J. Immunol. 186, 5706–5718 (2011).
Jensen, S. M., Potts, G. K., Ready, D. B. & Patterson, M. J. Specific MHC-I peptides are induced using PROTACs. Front. Immunol. 9, 2697 (2018).
Moser, S. C., Voerman, J. S. A., Buckley, D. L., Winter, G. E. & Schliehe, C. Acute pharmacologic degradation of a stable antigen enhances its direct presentation on MHC class I molecules. Front. Immunol. 8, 1920 (2017).
Gilbertson, B. & Subbarao, K. A new route to vaccines using PROTACs. Nat. Biotechnol. 40, 1328–1329 (2022).
Kavishna, R. et al. A single-shot vaccine approach for the universal influenza A vaccine candidate M2e. Proc. Natl Acad. Sci. USA 119, e2025607119 (2022).
Garber, K. The PROTAC gold rush. Nat. Biotechnol. 40, 12–16 (2022).
Salami, J. & Crews, C. M. Waste disposal—an attractive strategy for cancer therapy. Science 355, 1163–1167 (2017).
Ulmer, J. B., Valley, U. & Rappuoli, R. Vaccine manufacturing: challenges and solutions. Nat. Biotechnol. 24, 1377–1383 (2006).
Zhang, C. et al. PROTAR Vaccine 2.0 generates influenza vaccines by degrading multiple viral proteins. Nat. Chem. Biol. https://doi.org/10.1038/s41589-024-01813-z (2025)
Neumann, G. et al. Generation of influenza A viruses entirely from cloned cDNAs. Proc. Natl Acad. Sci. USA 96, 9345–9350 (1999).
Takada, A., Matsushita, S., Ninomiya, A., Kawaoka, Y. & Kida, H. Intranasal immunization with formalin-inactivated virus vaccine induces a broad spectrum of heterosubtypic immunity against influenza A virus infection in mice. Vaccine 21, 3212–3218 (2003).
Haredy, A. M. et al. An MDCK cell culture-derived formalin-inactivated influenza virus whole-virion vaccine from an influenza virus library confers cross-protective immunity by intranasal administration in mice. Clin. Vaccin. Immunol. 20, 998–1007 (2013).
Du, Y. et al. Genome-wide identification of interferon-sensitive mutations enables influenza vaccine design. Science 359, 290–296 (2018).
Broadbent, A. J., Santos, C. P., Godbout, R. A. & Subbarao, K. The temperature-sensitive and attenuation phenotypes conferred by mutations in the influenza virus PB2, PB1, and NP genes are influenced by the species of origin of the PB2 gene in reassortant viruses derived from influenza A/California/07/2009 and A/WSN/33 viruses. J. Virol. 88, 12339–12347 (2014).
Lei, C., Yang, J., Hu, J. & Sun, X. On the calculation of TCID50 for quantitation of virus infectivity. Virol. Sin. 36, 141–144 (2021).
Kastenschmidt, J. M. et al. Influenza vaccine format mediates distinct cellular and antibody responses in human immune organoids. Immunity 56, 1910–1926.e7 (2023).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587.e29 (2021).
Mathew, N. R. et al. Single-cell BCR and transcriptome analysis after influenza infection reveals spatiotemporal dynamics of antigen-specific B cells. Cell Rep. 35, 109286 (2021).
Hsieh, T. C., Ma, K. H. & Chao, A. iNEXT: an R package for interpolation and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 7, 1451–1456 (2016).
Luo, S. et al. Humanized V(D)J-rearranging and TdT-expressing mouse vaccine models with physiological HIV-1 broadly neutralizing antibody precursors. Proc. Natl Acad. Sci. USA 120, e2217883120 (2023).
Wang, L. et al. Generation of a live attenuated influenza vaccine that elicits broad protection in mice and ferrets. Cell Host Microbe 21, 334–343 (2017).
Acknowledgements
This study was supported by the National Key R&D Program of China (grant number 2024YFA0920000 to L.S.), Shenzhen Medical Research Fund (grant number B2303001 to L.S.), the National Natural Science Foundation of China (grant number 82273837 to L.S. and grant number 32101173 to Y.C.), Shenzhen Science and Technology Program (grant number JCYJ20220818101405011 to L.S.), the Strategic Priority Research Program of the Chinese Academy of Sciences (grant number XDB0480000 to C.L.), National Key R&D Program of China (grant number 2022YFC3400203 to Y.C.), China Postdoctoral Science Foundation (grant number 2022M723308 to J.L.), Guang Dong Basic and Applied Basic Research Foundation (grant number 2022A1515110061 to J.L.), and Key Laboratory of Quantitative Synthetic Biology, Shenzhen Institute of Synthetic Biology, Shenzhen Institutes of Advanced Technology, Chinese Academy of Sciences. We are grateful to the Shenzhen Infrastructure for Synthetic Biology for instrument support and technical assistance. We thank F. Yang, X. Shi, H. Li, W. Chen and C. Liu from the Shenzhen Institutes of Advanced Technology, Chinese Academy of Sciences, and X. Ou from the School of Life Sciences, Southern University of Science and Technology, for helpful suggestions.
Author information
Authors and Affiliations
Contributions
L.S. conceived this study. L.S., J.S., J.L., Q.S., J. Hou, C.Z., H.B. and X.A. designed the experiments and analysed the data. J.S., J.L., J.H. and C.Z. performed cell culture experiments and collected data with assistance from Q.S., Z.W., J. Hao, P.W., Q.Z., Z.L., L.L., Q.W., Y.Z., Y. Su, C.L., X. Xi and J.X. Q.S. performed animal experiments and collected data with assistance from J.L., J.S., C.Z., P.W. and L.L.; H.B., X.A., B.X., T.F., L.Y., Y. Sun, X. Xie, J.X. and R.P. performed human lung airway-on-a-chip experiments and collected data. J.L., Q.S., A.Y.Y., H.B. and H.H. performed RNA-seq, scRNA-seq, scBCR-seq and data analysis. F.Q., Y.C., J.X., X.L., R.D., R.P., L.Z., D.Z. and G.C. provided relevant materials, technical assistance and insightful input in terms of experiment design, data analysis and writing. L.S. wrote the paper, which all authors edited.
Corresponding author
Ethics declarations
Competing interests
L.S., J.S., J.L. and Q.S. are inventors on relevant patent applications held by Shenzhen Institutes of Advanced Technology, Chinese Academy of Sciences. The other authors declare no competing interests.
Peer review
Peer review information
Nature Microbiology thanks Alessio Ciulli and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Systematic investigation on the propagation competence of PROTAR viruses in MDCK-TEVp and conventional MDCK.2 cells.
MDCK-TEVp cells or MDCK.2 cells were infected with indicated viruses (MOI = 0.001) for 48 h and supernatants were collected for quantification of viral titers by TCID50 assay. The factors of decrease in viral titers of PROTAR viruses in MDCK.2 cells relative to those in MDCK-TEVp cells are indicated in the graph. The dotted line indicates the detection limit (102 TCID50/mL). Data are shown as means ± s.d. The experiments were independently performed two times.
Extended Data Fig. 2 Western blots showing co-IP of E3 ubiquitin ligases with the PTD-tagged viral M1 proteins but not the mutated PTD-tagged viral M1 proteins.
HEK293T cells were transfected with constructs expressing Flag- or HA-tagged E3 ubiquitin ligases and either PTD-tagged viral M1 proteins or mutated PTD-tagged viral M1 proteins and collected 36 h post-transfection for co-IP assay (n = 3). WCL, whole-cell lysate.
Extended Data Fig. 3 Western blots showing ubiquitination of PTD-tagged viral M1 proteins.
HEK293T cells were co-transfected with constructs expressing HA- or Flag-tagged ubiquitin, Flag- or HA-tagged E3 ubiquitin ligases, and PTD-tagged viral M1 proteins or mutated PTD-tagged viral M1 proteins. The cells were treated with 10 μM MG-132 for 6 hours prior to being harvested at 36 h post-transfection. M1 proteins were immunoprecipitated for detection of the levels of ubiquitination (n = 3).
Extended Data Fig. 4 PTD-mediated proteasomal degradation of viral M1 protein of PROTAR viruses is dependent on their respective E3 ubiquitin ligase.
Conventional cells were transfected with indicated small interfering RNAs (siRNAs) (20 nM) or short hairpin RNAs (shRNAs) (1 µg/mL) for knockdown of each of the E3 ubiquitin ligases or scrambled RNA control (Ctrl RNA), and 24 h later infected with indicated viruses (MOI = 0.1). 48 h after infection, cells were collected for detection of the levels of influenza M1 protein by western blotting (n = 3).
Extended Data Fig. 5 PTD-mediated proteasomal degradation of viral M1 protein of PROTAR viruses is E3 ubiquitin ligase-dependent.
a, Inhibition of Cullin-RING E3 ubiquitin ligases restored the levels of PTD-tagged viral M1 proteins. HEK293T cells were transfected with constructs expressing indicated proteins for 6 h and cultured in the presence or absence of 1 μM MLN4924 for another 18 hours. Cells were harvested and subjected to detection of M1 proteins by western blotting (n = 3). b, Inhibition of E3 ubiquitin ligases by specific inhibitors restored the levels of PTD-tagged viral M1 proteins. HEK293T cells were transfected with constructs expressing indicated proteins for 48 h and cultured in the presence or absence of indicated inhibitors for another 6 hours. Cells were harvested and subjected to detection of M1 proteins by western blotting (n = 3).
Extended Data Fig. 6 Humoral immune responses induced by PROTAR vaccines in adult mice.
a-b, Titers of NT (a) and HI (b) antibodies in sera of C57BL/6 J mice at day 21 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group). Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test; black P values indicate the statistical comparison to Vehicle group; orange P values indicate the statistical comparison to IIV group; red P values indicate the statistical comparison to M1-PTD. c-d, Titers of IgG antibodies against viral surface protein HA (c) and internal conserved protein NP (d) in sera of C57BL/6 J mice at day 21 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group). Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test; black P values indicate the statistical comparison to Vehicle group; orange P values indicate the statistical comparison to IIV group; blue P values indicate the statistical comparison to CAIV group; red P values indicate the statistical comparison to M1-PTD.
Extended Data Fig. 7 Mucosal immune responses induced by PROTAR vaccines in adult mice.
Titers of IgA antibodies against whole influenza virions (a), HA (b), NA (c), and NP (d) in bronchoalveolar lavage (BAL) of C57BL/6 J mice at day 21 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group). Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test; black P values indicate the statistical comparison to Vehicle group; orange P values indicate the statistical comparison to IIV group; blue P values indicate the statistical comparison to CAIV group; red P values indicate the statistical comparison to M1-PTD.
Extended Data Fig. 8 T cellular immune responses induced by PROTAR vaccines in adult mice.
a, Viral M1 antigen peptide-specific T cell responses in lungs of C57BL/6 J mice at day 7 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group), measured by enzyme-linked immunospot (ELISpot) assay. M1128-135 (MGLIYNRM) antigen peptide was used as the stimuli. Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test; black P values indicate the statistical comparison to Vehicle group; orange P values indicate the statistical comparison to IIV group; blue P values indicate the statistical comparison to CAIV group; red P values indicate the statistical comparison to M1-PTD. b-c, Viral NP antigen peptide-specific T cell responses in lungs (b) and spleens (c) of C57BL/6 J mice at day 7 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group), measured by enzyme-linked immunospot (ELISpot) assay. NP366-374 (ASNENMETME) antigen peptide was used as the stimuli. Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test; black P values indicate the statistical comparison to Vehicle group; orange P values indicate the statistical comparison to IIV group; blue P values indicate the statistical comparison to CAIV group; red P values indicate the statistical comparison to M1-PTD.
Extended Data Fig. 9 Analysis of viral M1 antigen presentation of PROTAR vaccines.
a, Viral M1 antigen presentation of PROTAR vaccines, IIV, CAIV, and WT virus. Raw264.7 cells were infected with indicated vaccines or viruses (MOI = 0.1) for 6 hours and detected for M1 antigen presentation on the surface of Raw264.7 cells by anti-M1 antibody (left) or an anti-M1 peptide (MGLIYNRM) antibody (right) using flow cytometry. Data are shown as means ± s.d.; n = 3 biological replicates; one-way ANOVA with Dunnett’s multiple comparisons test; black P values indicate the statistical comparison to WT group; orange P values indicate the statistical comparison to IIV group; blue P values indicate the statistical comparison to CAIV group; red P values indicate the statistical comparison to M1-PTD. b, Graphs showing the relevance between PTD-mediated antigen presentation and the levels of PROTAR vaccine-induced immune responses, including IgG antibody responses (top left), IgA antibody responses (top middle and right), T cellular responses against M1 antigen in lungs (bottom left), T cellular responses against NP antigen in lungs (bottom middle), and T cellular responses against NP in spleens (bottom right) (n = 5 mice per group). The same data for M1 antigen presentation are used in all the graphs.
Extended Data Fig. 10 Characterization of B cell responses induced by PROTAR vaccines in adult mice.
a, Graphs showing numbers of HA- or NP-binding GC B cells per 20,000 cells in spleens of C57BL/6 J mice at day 14 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group). Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test. b, Graphs showing numbers of HA- or NP-binding memory B cells per 20,000 cells in spleens of C57BL/6 J mice at day 14 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group). Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test. c, Graphs showing numbers of HA- or NP-binding plasma cells per 20,000 cells in spleens of C57BL/6 J mice at day 14 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group). Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test. d. Graphs showing numbers of influenza antigens (HA or NP)-specific IgG antibody-secreting plasma cells in spleens or lungs of C57BL/6 J mice at day 14 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group). Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test. e, Graphs showing numbers of influenza antigens (HA or NP)-specific memory B cells in spleens or lungs of C57BL/6 J mice at day 14 post-vaccination with 105 TCID50 of indicated vaccines (n = 5 mice per group). Memory B cells from mouse spleens or lungs were stimulated to proliferate and differentiate into antibody-secreting cells by R848 (1 µg/mL) and recombinant mouse (rm)IL-2 (10 ng/mL) in vitro, and levels were subsequently measured by B-cell ELISpot assay. Data are shown as means ± s.d.; one-way ANOVA with Dunnett’s multiple comparisons test. f, Immunofluorescence images showing GC formation in lungs (left) and spleens (right) of C57BL/6 J mice at day 14 post-vaccination with 105 TCID50 PTDβ-TrCP. Representative images are shown; n = 3 biological replicates. Green, GL7; Red, B220; Blue, DAPI-stained nuclei. Scale bar, 100 µm.
Supplementary information
Supplementary Information
Supplementary Figs. 1–12, Tables 1–5 and source data for Supplementary Fig. 4.
Source Data Supplementary Fig. 1
Statistical source data.
Source Data Supplementary Fig. 5
Statistical source data.
Source Data Supplementary Fig. 6
Statistical source data.
Source Data Supplementary Fig. 7
Statistical source data.
Source Data Supplementary Fig. 10
Statistical source data.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Unprocessed western blots.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Unprocessed western blots.
Source Data Extended Data Fig. 3
Unprocessed western blots.
Source Data Extended Data Fig. 4
Unprocessed western blots.
Source Data Extended Data Fig. 5
Unprocessed western blots.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 9
Statistical source data.
Source Data Extended Data Fig. 10
Statistical source data.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Shen, J., Li, J., Shen, Q. et al. Proteolysis-targeting influenza vaccine strains induce broad-spectrum immunity and in vivo protection. Nat Microbiol 10, 431–447 (2025). https://doi.org/10.1038/s41564-024-01908-2
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41564-024-01908-2
This article is cited by
-
Synthetic biology-inspired development of live attenuated influenza vaccines
npj Vaccines (2025)
-
Comparison of efficacy, safety, immune response of dNS1 LAIV and cold-adapted LAIV in a mouse model
npj Vaccines (2025)
-
Live vaccine development through targeted protein degradation
Nature Reviews Immunology (2025)
-
PROTAR Vaccine 2.0 generates influenza vaccines by degrading multiple viral proteins
Nature Chemical Biology (2025)
-
Harnessing molecular proximity for antiviral innovations: advances in proximity-inducing modalities against viral infections
Science China Chemistry (2025)
