
Abstract
Innate lymphoid cells (ILCs) are tissue-resident lymphocytes specialized in cytokine secretion that lack antigen-specific receptors. The contribution of ILCs to antiviral mucosal immunity in humans, particularly in the female genital tract (FGT), remains unexplored. Here we resolved human FGT ILC diversity by spectral flow cytometry and CITE-seq, spatial location within genital anatomical regions using ChipCytometry, and determined homeostatic function and antiviral responses. We uncovered spatial and functional specializations of genital ILC subsets under homeostasis, with compartmentalized age-related and pregnancy-related changes. CD161 expression differentially discriminated ILC subsets preloaded with cytokines at steady state. We identified a unique NKp44+CCR6+ ILC3 subset in the endometrium that actively degranulated at homeostasis and was located in lymphoid aggregates surrounded by B cells and T cells. By contrast, ILC1s were found scattered, enriched in the ectocervix and located close to the epithelium. Following in vitro HIV stimulation, genital ILCs displayed rapid subset-specific antiviral responses. These findings reveal distinct tissue and subset-specific features of FGT ILCs and their capacity to immediately respond to viral stimuli, providing a foundation for future studies to determine the potential role of ILCs in mucosal immune protection in the FGT.
Similar content being viewed by others

Main
Women acquire human immunodeficiency virus (HIV) primarily through sexual transmission1. However, under non-inflammatory conditions, HIV transmission rates per sexual act are low (0.08%) but as high as 2.65% in the context of sexually transmitted infections2. Protective mechanisms in the FGT include intact epithelial barriers, mucus, secretions and innate immune cells3,4,5,6,7,8. Despite this knowledge, we still lack an understanding of the initial events of infection and how to enhance FGT mucosal protection. This is in part because initial mucosal host–pathogen interactions that determine whether infection is prevented or established happen very quickly (within minutes) and cannot be captured in vivo. To develop effective preventative methods, it is imperative to dissect the early events of mucosal HIV acquisition and how the different innate immune cells respond to HIV in the female genital mucosa, one of the main portals of entry for HIV.
ILCs are tissue-resident lymphocytes that lack antigen-specific receptors and specialize in cytokine secretion. Helper ILCs are grouped into three main populations (ILC1s, ILC2s and ILC3s) that functionally resemble TH1, TH2 and TH17 CD4+ helper T cells9. ILC1s produce interferon-γ (IFNγ); ILC2s secrete IL-5, IL-13 and IL-9; and ILC3s produce IL-22 and IL-17 in response to stimuli9. Murine ILCs contribute to barrier integrity and mucosal protection against infection10, but ILCs in human mucosal surfaces and their contribution to mucosal protection remain largely uncharacterized11,12,13.
Tissue-specific adaptations, lack of lineage markers and overall rare presence make human ILC characterization difficult and highlight the need to define human ILC subsets in their tissue of residence through combined molecular analyses11,14,15,16. Human ILCs are actively studied in the tonsils14,17,18,19, skin20, gastrointestinal13 and respiratory tracts12, but remain largely uncharacterized in the FGT. Helper ILCs have been described to a limited extent in the human endometrium (EM) and decidua during pregnancy21,22, but their role in mucosal protection throughout the non-pregnant FGT remains unknown. Studies of blood, gut and tonsils from patients infected with HIV have described ILC alterations in the context of chronic infection18,23,24; however, the genital mucosa, a major site for HIV infection, has not been investigated25. Importantly, the role that ILCs have in mucosal HIV pathogenesis immediately after exposure, and whether they can respond to HIV stimulation, is completely unknown.
The immune system in the FGT is different from other mucosal surfaces, as it is uniquely conditioned and regulated by sex hormones to perform reproductive functions and protect against pathogens7. Each anatomical region of the FGT exhibits discrete histological, immunological and functional traits. The lower FGT, consisting of the ectocervix (ECT) and vagina, is covered by stratified squamous epithelial cells, populated by commensal microbes and serves as the area of semen reception7. By contrast, the upper FGT, which includes the uterine endocervix (END) and endometrium (EM), is lined with a single layer of columnar epithelium, has fewer commensal microbes and is specialized to facilitate embryo implantation and pregnancy7.
Previously, we demonstrated immune compartmentalization of T cell populations throughout the FGT, including differences in TH17 CD4+ T cell distribution and susceptibility to HIV infection26,27,28,29, emphasizing the importance of studying the entire FGT. Defining how—throughout the FGT—ILCs maintain tissue homeostasis in a mucosa dually tasked with pathogen protection and reproduction, along with their response to HIV, will fill a gap in knowledge of mucosal defense against pathogens and open new avenues for therapeutic interventions.
Here we used a multi-omics approach with high-dimensional flow cytometry, CITE-seq and ChipCytometry to deeply characterize the distribution and spatial location of helper ILCs throughout the human FGT, including the EM, END and ECT. Moreover, we defined the ability of ILCs to respond to HIV challenge in a subset-specific manner, suggesting a role for genital ILCs in mucosal antiviral responses.
Results
ILCs reside in the female genital mucosa and are armed with antiviral cytokines at homeostasis
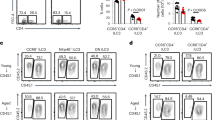
ILC presence and subset distribution under homeostatic conditions throughout the FGT remain uncharacterized. To address this knowledge gap, we assessed ILC subsets using high-dimensional spectral flow cytometry. Genital ILCs were identified as CD45+ cells that lacked canonical lineage markers (CD3, CD19, CD11b, CD14, CD16, CD66b, CD11c) and expressed CD127 (IL-7R) (Fig. 1a), according to established definitions in other human mucosal surfaces. Our gating strategy excluded canonical natural killer (NK) cells, determined by lack of expression of NKp46 (CD335) and CD94 on ILCs, and lack of CD127 expression on NK cells9 (Extended Data Fig. 1a,b). Additionally, NK cells expressed high levels of CD56 and CD94, whereas helper ILCs expressed low levels of CD56 throughout the FGT (Extended Data Fig. 1c,d). Our strategy excluded other rare cell types that may contaminate the ILC gate. We did not detect CD203c expression (a marker of mast cells and basophils30), CD34+ progenitor cells or CD123+ plasmacytoid dendritic cells (Extended Data Fig. 1e–g).

a, Representative gating strategy to identify genital lineage-negative, CD127+ ILCs. SSC-A, side scatter area; FSC-A, forward scatter area; FSC-H, forward scatter height; FMO, fluorescence minus one. b, Quantification of ILC percentage in EM (n = 26), END (n = 18) and ECT (n = 16) per total mononuclear cells and ILC number per gram of tissue (EM, n = 22; END, n = 17; ECT, n = 15 tissues). c, Contour plot showing ILC subset gating strategy. d, Comparison of percentage of ILC subsets in total ILCs across three tissues (n = 56). e, Pie charts indicating the percentage of patients with 0, <5% or >5% ILC2s in total ILCs (EM, n = 25; END, n = 16; ECT, n = 15). f, Contour plot showing ILC subsets along with matching histograms of transcription factor expression within the gated ILC subsets. g, Representative contour plots indicating percentage of ILC1s, ILC2s and ILC3s positive for T-bet, GATA-3 and RORyt, respectively. The FMO strategy was used to determine positive gates. Quantification of multiple donors (right): T-bet (n = 6), GATA-3 (n = 4) and RORyt (n = 4). h,i, Contour plot and quantification of IFNγ (n = 45 tissues: EM, n = 18; END, n = 10; ECT, n = 17) (h) and IL-22 expression in genital ILCs (n = 36 tissues: EM, n = 16; END, n = 9; ECT, n = 11) (i). j,k, Histogram of IFNγ (j) and IL-22 (k) expression in ILCs, CD3+ T cells, CD56hi CD127− NK cells and FMO. Each dot represents a biological replicate. Horizontal lines represent the median ± interquartile range (IQR). Statistical analysis was performed using the non-parametric Kruskal–Wallis test with Dunn’s post-test correction for multiple comparisons.
ILCs represented approximately 0.52% of the total mononuclear immune cells, with no differences between tissues, and the number of ILCs per gram of tissue was significantly higher in EM than in ECT (Fig. 1b).
Next, we quantified genital ILC subsets, according to the expression of CD294 (ILC2s), CD117 (ILC3s) or double-negative populations (ILC1s) as previously described10 (Fig. 1c). ILC1s and ILC3s were the main subsets present but showed compartmentalization, with ILC1s enriched in the END and ECT, while ILC3s were enriched within the EM (Fig. 1d). ILC2s displayed very low frequencies throughout the FGT (Fig. 1d), with more than half of the patients having less than 5% or no ILC2s (Fig. 1e). Genital T cells (CD3+) from the same samples displayed high CD294 expression (Extended Data Fig. 1h), demonstrating that the lack of CD294 expression on ILCs was a result of low ILC2 presence in genital tissues and not caused by faulty antibody staining. ILC3s displayed higher CD127 expression than ILC1s (Extended Data Fig. 1i).
Next, we assessed canonical transcription factor expression: T-bet (ILC1s), GATA-3 (ILC2s) and RORyt (ILC3s)10. ILC1s (CD294−CD117−) expressed T-bet, ILC2s (CD294+) expressed GATA-3 and ILC3s (CD117+) expressed RORyt (Fig. 1f). Approximately 80% of ILC1s were T-bet+, 95% of ILC2s were GATA-3+ and 98% of ILC3s were RORyt+, consistently across samples (Fig. 1g). Our transcription factor expression patterns identified a small population of ILC1-like cells that do not express T-bet (Fig. 1f,g), similar to recent findings in other human mucosal surfaces19.
Lastly, we investigated cytokine production to define the functional status of ILCs in the genital mucosa under homeostatic conditions. In the absence of stimulation, intracellular staining showed IFNγ production by genital ILC1s (Fig. 1h; CD117−) and IL-22 by ILC3s (Fig. 1i; CD117+). IFNγ+ ILC1s and IL-22+ ILC3s represented about 10% and 45%, respectively, of each ILC subset across all tissues (Fig. 1h,i). Given that most immune cells do not constitutively produce IFNγ or IL-22, we compared cytokine expression in ILCs to unstimulated NK cells and T cells from the same tissues. Interestingly, we observed that genital ILCs were the main producers of IFNγ and IL-22 under homeostatic conditions (Fig. 1j,k). Of note, genital ILCs did not constitutively produce IL-17, IL-5, IL-13 or NK-related cytolytic molecules such as Granulysin (Extended Data Fig. 1j,k).
In summary, genital ILC subsets are compartmentalized and loaded with specific antiviral cytokines under homeostatic conditions, suggesting potential roles in first-line antiviral mucosal protection.
Unbiased clustering reveals discriminatory markers of genital ILC subpopulations and markers of tissue residency
To further uncover discriminatory markers that define functional ILC subpopulations, we performed unbiased clustering of high-dimensional flow cytometry data.
Genital ILCs clustered closely based on tissue type (Fig. 2a, upper panel). Individual clustering per patient is shown in Extended Data Fig. 2a. ILCs from the EM (Fig. 2a,b, red) clustered more independently than ILCs from the END and ECT (Fig. 2a,b, blue and green), suggesting that endometrial ILCs are phenotypically more distinct than cervical ILCs.

a, opt-SNE plots on concatenated total ILCs from three matched patients’ EM, END and ECT samples (top). FlowSOM clustering analysis overlayed onto the opt-SNE plot with ten distinct clusters (bottom). b, opt-SNE plot of total ILCs from three patients split by anatomical region. c, opt-SNE plot of total ILCs with individual protein expression overlayed. The color determines the intensity of expression (blue, low; red, high). d, Contour plot of CD103 and CD69 expression in blood and genital ILCs. e,f, Quantification of the percentage of CD69 (e) and CD103 (f) expressing genital ILCs. g, Quantification of the percentage of CD103+ ILCs by subset across tissues (n = 56 tissues). h, Representative histogram of CCR6 expression in each ILC subset. i, Quantification of CCR6 expression in ILC3s. j, Contour plot of CD103 and CCR6 expression in ILC3s (left) with CCR6 FMO. Quantification of ILC3 subsets in all tissues (right). k, Pie graph of the percentage of ILC3 subsets in total ILC3s. l, Percentage of CD161+ ILCs in each tissue. m, Contour plot of CD161 expression. n, Percentage of CD161+ ILC1s and ILC3s in the EM, END and ECT. o, Contour plot of CD161 and CD103 expression in ILC1s. p, Percentage of CD161+ ILC1s in CD103+ and CD103− ILC1s. q, Contour plot of CD161 and CD103 expression in ILC3s. r, Percentage of CD161+ ILC3s in CCR6+CD103+, CCR6+CD103− and CCR6− ILC3s. Each dot represents a biological replicate; n = 61 tissues. Horizontal lines represent the median ± IQR. Statistical analysis was performed using the non-parametric Kruskal–Wallis test with Dunn’s post hoc test correction for multiple comparisons.
Next, to uncover genital ILC subpopulations, we used unsupervised clustering analysis (FlowSOM) to perform multivariate clustering of cells based on self-organized maps. ILCs were grouped into ten clusters that were overlayed on the opt-SNE plots (Fig. 2a, lower panel). This analysis identified tissue-dependent clusters, such as cluster 7 or 9, while others were intermixed (cluster 5 or 3) (Fig. 2a). To identify key drivers of clustering, we overlaid individual proteins on the opt-SNE plots (Fig. 2c). Most ILCs expressed high levels of CD69, a marker for tissue residency. Analysis of protein expression in each cluster identified surface markers that revealed anatomical differences, like CD161 and CXCR4 (Fig. 2c), and initial assignment of clusters to specific ILC subsets (Extended Data Fig. 2b). We also identified discriminatory markers for exploration of specific ILC clusters, mainly CD103, CCR6, CD161 and CD107a (Fig. 2a–c and Extended Data Fig. 2c). Importantly, stratification of functional markers (IFNy, IL-22 and CD107a) per individual patient and by tissue revealed tissue-specific differences and patient-to-patient variation (Extended Data Fig. 2d), indicating the need for further evaluation in a larger cohort.
Therefore, we validated our findings with a larger number of patients (n = 32). The majority of genital ILCs expressed CD69 (Fig. 2d), with no differences between anatomical sites (Fig. 2e). Additionally, a proportion of CD69+ ILCs co-expressed CD103 (Fig. 2d) with no differences between anatomical sites (Fig. 2f) or ILC subsets (ILC1, ILC3) (Fig. 2g). ILC2s were excluded from these analyses because of low cell number and absence of this population in a large proportion of patients (Fig. 1c,d).
Our unbiased clustering analysis indicated high levels of co-expression between CD103 and CCR6, a chemokine receptor for CCL20, which is produced by genital epithelial cells31. CCR6 was highly expressed on ILC3s (Fig. 2h) and preferential to the EM (Fig. 2i). The majority of CD103+ ILC3s co-expressed CCR6 (Fig. 2j), allowing the discrimination of three unique ILC3 subpopulations based on CD103 and CCR6 expression: CCR6−, CCR6+CD103− and CCR6+CD103+ ILC3s (Fig. 2j). A representative dot plot of CCR6 versus CD45 expression and corresponding fluorescence-minus-one control is shown to clarify the gating strategy (Extended Data Fig. 2e). Quantification of each ILC3 subset across the FGT showed enrichment of CCR6− in ECT and CCR6+CD103− in the EM, while the CCR6+CD103+ population remained constant across tissues (Fig. 2k).
CD161, a site-specific discriminatory marker identified in the opt-SNE plot, showed significant differential expression between endometrial and cervical ILCs (Fig. 2l,m) but similar expression between ILC1s and ILC3s (Fig. 2n). Furthermore, CD161 was preferentially co-expressed on CD103+ ILC1s and ILC3s (Fig. 2o–q), particularly on the CD103+CCR6+ ILC3 subpopulations in the END and ECT (Fig. 2r).
CD161 and NKp44 discriminate functional ILC subsets at homeostasis
We next investigated potential associations between phenotypical markers (CD103, CD161 and CCR6) and functional specialization of ILC subsets. IFNγ production by ILC1s was independent of CD103 expression (Fig. 3a and Extended Data Fig. 3a), but significantly enhanced in CD161− ILC1s (Fig. 3b and Extended Data Fig. 3b). IL-22 production was highest in CCR6+CD103+ ILC3s compared to CCR6+CD103− and CCR6− ILC3s (Fig. 3c and Extended Data Fig. 3c). Furthermore, CD161+ ILC3s displayed enhanced IL-22 production (Fig. 3d and Extended Data Fig. 3d). These data suggest that CD161 is a relevant marker to discriminate ILC subsets with high cytokine production at homeostasis.

a, Representative histogram (left) and contour plot (middle) of IFNγ expression in CD103+ and CD103− ILC1s with quantification of percentage of IFNγ+ ILC1s (right) (n = 30). b, Histogram and contour plot of IFNγ expression in CD161+ and CD161− ILC1s (left) (n = 30). Quantification of IFNγ+CD103− and CD103+ ILC1s based on CD161 expression (right). c, Histogram and contour plot of IL-22 expression in ILC3 subsets (left) with quantification of percentage of IL-22+ ILC3 subsets (right) (n = 30). d, Histogram and contour plot of IL-22 expression in CD161+ and CD161− ILC3s. Quantification of IL-22+CCR6+CD103+ and CCR6+CD103− ILC3s based on CD161 expression (right) (n = 30). e, Representative contour plot of CD107a expression in FMO (left) and EM (right) ILCs. f, Histogram of CD107a expression in ILCs, NK cells and T cells. g, Contour plot of CD107a and CD117 in total ILCs. h, Contour plot of CD103 and CD107a in ILC3s. i, Histogram and contour plot of CD107a expression in all ILC subsets. j, Percentage of CD107a+ cells by ILC3 subsets. k, Percentage of CD107a+CCR6+CD103− ILC3s by tissue (n = 24 tissues). l, Contour plot of CD161 and CD107a expression (left) and mean fluorescence intensity (MFI) comparison between CD107a+ and CD107a− in n = 12 EM samples. m, Contour plot of NKp44 and CD117 (left) and percentage of NKp44+ ILC3s by tissue (right) (n = 17 tissues: EM, n = 7; END, n = 5; ECT, n = 5). n, Representative histogram of NKp44 expression in ILC subsets (left). Percentage of NKp44+ ILC3s by ILC3 subset (right) (n = 9 tissues). o, Contour plot of CD107a and NKp44 expression in EM (top) and END (middle) ILC3s. Percentage of NKp44+ ILC3s that are CD107a+ (bottom) (n = 10 tissues: EM, n = 5; END and ECT, n = 5). p, Contour plots representing IL-22 secretion during PMA stimulation for 1 h in NKp44+/− ILC3s. Graph on the right represents the percentage of ILC3s positive for IL-22 secretion with and without stimulation (n = 6 tissues). q, Representative contour plots of CD107a expression in genital NKp44+/− ILC3s with and without PMA stimulation. Graph on the right represents the percentage of ILC3s positive for CD107a with and without stimulation (n = 6). Each dot represents a biological replicate. Horizontal lines represent the median ± IQR. Statistical analysis was performed using the non-parametric Kruskal–Wallis test with Dunn’s post hoc test correction for multiple comparisons.
We next investigated ILC degranulation (CD107a expression) as an additional functional parameter detected in our unbiased analysis (Fig. 2c). Although CD107a is normally expressed on the cell surface following stimulation, we detected CD107a expression on a high percentage of genital ILCs in the absence of in vitro stimulation (Fig. 3e). Importantly, CD107a expression was limited to ILCs, while genital T cells and NK cells from the same unstimulated samples did not express CD107a (Fig. 3f). Further analysis demonstrated that degranulation was specific for ILC3s (CD117+; Fig. 3g), and particularly CCR6+CD103− ILC3s (Fig. 3h–j and Extended Data Fig. 3e). However, degranulation occurred primarily in the EM, with little to no CD107a expression in the END and ECT (Fig. 3k). Given that CD161 identified ILC3s with high intracellular IL-22 (Fig. 3d), we assessed whether CD161 expression was also related to degranulation. However, endometrial CD107a+ ILC3s expressed significantly lower levels of CD161 (Fig. 3l).
As we observed a wide range of IL-22, IFNy and CD107a expression between patients, we investigated whether cytokine production and degranulation could identify patients with ‘highly functional’ ILCs. We performed correlations between IL-22, IFNγ and CD107a across patients; however, we did not detect any correlations (Extended Data Fig. 3f), suggesting that ILC subsets function independently of each other under homeostatic conditions.
To better define the ILC3 degranulating population, we evaluated NKp44 expression, previously described in intestinal32, endometrial and decidual ILCs21,22. NKp44 was exclusively expressed on ILC3s (CD117+) and was present throughout the FGT, although significantly enriched in the EM (Fig. 3m). NKp44 expression was specific to CCR6+CD103− ILC3s (Fig. 3n), the same subset undergoing degranulation (Fig. 3i,j). Consistently, NKp44+ ILC3s co-expressed CD107a and were found primarily at the EM (Fig. 3o), indicating ongoing endometrial ILC3 degranulation at homeostasis. To explore whether the NKp44+ and NKp44− ILC3 subsets were able to rapidly secrete the IL-22 stored intracellularly, and degranulate in response to stimulation, we performed a flow-cytometry-based IL-22 secretion assay. A small percentage of ILC3s secreted IL-22 under resting conditions (Fig. 3p, control), but IL-22 secretion was significantly increased in response to phorbol 12-myristate-13-acetate (PMA) stimulation (Fig. 3p, PMA). Moreover, NKp44− ILC3s secreted IL-22 in response to PMA stimulation (Fig. 3p), while NKp44+ ILC3s significantly upregulated CD107a expression after stimulation (Fig. 3q), indicating different functionalities between these two ILC3 subsets.
NKp44+ ILC3s reside in lymphoid aggregates within the EM
Our data indicate the presence of multiple ILC subpopulations within the tissue, each with distinct functional phenotypes. Given that ILCs are known to interact with the tissue environment and regulate other cell types, we investigated the spatial location and cell interactions of genital ILCs to understand the functional consequences for these distinct phenotypes. As ILCs lack canonical lineage markers and are difficult to accurately identify with conventional confocal microscopy, we used ChipCytometry to achieve multiplexed spatial fluorescence microscopy. Building on our flow cytometry data, we identified ILCs as lineage-negative (CD3, CD20, CD11c, CD14, CD16, CD66b) CD127+ cells (Extended Data Fig. 4a) and visualized functional ILC subpopulations through the expression of CD117, CCR6, CD103, CD161 and NKp44 (Extended Data Fig. 4a).
In the EM, we identified ILCs positioned within large clusters of CD45+ cells located near endometrial epithelial glands (Fig. 4a). These cell aggregates consisted largely of lymphocytes, including CD3+ T cells and CD20+ B cells. Within the lymphoid aggregates, we exclusively found ILC3s (CD117+) co-expressing CCR6 and NKp44 (Fig. 4a,b). We identified both CD161+ and CD161− NKp44+ ILC3s in the aggregates, closely interacting with CD4+ and CD4− T cells (CD3+), and B cells (CD20+) (Fig. 4b). Additionally, we observed clusters of NKp44+ ILC3s (CD161+ and CD161−) localizing to the edge of the glands present in the EM, interacting closely with epithelial cells (PanCk+) (Fig. 4c). We did not find NKp44− ILC3s within the aggregates (Fig. 4a–c). We identified CCR6+NKp44− ILC3s positioned close to endometrial epithelial glands, interacting with CD45− stromal cells but not in contact with T cells or B cells (Fig. 4d). To quantify the NKp44+ and NKp44− ILC3 subsets, we selected regions of interest with ILC3s that contained lymphoid aggregates and those that did not (Extended Data Fig. 4b). In these regions, we counted the number of NKp44+/− ILC3s per area. The majority of NKp44− ILC3s were located outside of lymphoid aggregates, while almost all NKp44+ ILC3s were found in the lymphoid aggregates (Fig. 4e).

a, Representative image of lymphoid aggregates in the EM (top) and zoomed in (bottom). PanCK (white) identifies epithelial cells; DAPI (blue) identifies nuclei. b, Representative image of NKp44+ ILC3s at 10 µm. Images are separated by channels. Red arrows denote CD161+ ILC3s; white arrows denote CD161− ILC3s. c, Zoomed-in image of NKp44+ ILC3s in the aggregate near PanCK+ cells. d, NKp44− ILC3 in the EM. e, Quantification of NKp44+ and NKp44− ILC3s per area (log10 scale) in regions of interest (ROIs) that contained ILC3s: lymphoid aggregates (LA; n = 5 ROIs) and non-lymphoid aggregates (non-LA; n = 15 ROIs). Horizontal lines represent the median ± IQR. f, CD103−CD161+ ILC1s in the ECT. g, CD103+CD161− ILC1s in the END near the epithelium; n = 3 donors, with nine tissues total (EM, END and ECT for each). h, Number of CD161+, CD161− and CD103+ ILC1s per area (log10 scale) in ROIs containing ILCs: intraepithelial (n = 3 ROIs including the entirety of the epithelium from three patients), subepithelial (n = 5 ROIs) and deeper tissue (n = 7 ROIs). Each dot represents one ROI. ROIs from three patients were used. Horizontal lines represent the median ± IQR. Kruskal–Wallis test with Dunn’s post hoc test correction for multiple comparisons.
In the ECT, we primarily detected ILC1s (Fig. 4f,g). In contrast to the EM, the ILCs identified in the ECT did not cluster together and were not located within cell aggregates. CD161+ and CD161− ILC1s were present close to the stratified ectocervical epithelium and preferentially interacted with T cells (Fig. 4f,g). CD103+ ILC1s were located closer to the basal layer of the stratified epithelium (Fig. 4g). To quantify ILC1 subsets (CD161+, CD161− and CD103+) in the ECT, we selected areas of interest based on tissue location relative to the epithelium (intraepithelial, subepithelial and deeper tissue; Extended Data Fig. 4c). ILC1s were located in the subepithelial or deeper tissue compartments, while no ILC1s were found within the epithelium (Fig. 4h). Furthermore, CD103+ and CD161− ILC1s were preferentially located in the subepithelial compartment, while CD161+ ILC1s were more abundant deeper in the tissue and did not express CD103 (Fig. 4h).
Our data indicate that the functional markers CD103, CD161 and NKp44 also associate with ILC spatial location and potential cell interactions.
Genital ILC subsets are regulated by age and prior pregnancies
Given that we observed anatomical compartmentalization and high donor-to-donor variability in genital ILC distribution and phenotype, we investigated whether ILCs are modified according to age and reproductive parameters. To determine age-associated effects on ILCs, we performed correlation analyses. As women age, the percentage of ILCs in total mononuclear cells decreases in ECT but remains unchanged in EM and END (Fig. 5a). However, ILC subset analysis showed an age-dependent switch in subset distribution in the EM specifically, with a reduction in ILC1 percentage (Fig. 5b) concomitant with an ILC3 increase (Fig. 5c), while ILC subset distribution in the END and ECT was not affected by age (Fig. 5b,c). Given these differential effects in the EM, we investigated whether prior pregnancies affected ILC frequency. First, we compared donors who were never pregnant to donors who had at least one pregnancy, but no differences were found (Fig. 5d). However, we observed that time after last pregnancy positively correlated with ILC percentage in the EM. Donors who were recently pregnant had lower numbers of endometrial ILCs than donors not recently pregnant (Fig. 5e). We observed no trend with donors who were never pregnant. Importantly, the effect of time after last pregnancy on ILC frequency was selective to the EM (Fig. 5e, top graph).

a, Spearman correlation of percentage of ILCs in mononuclear cells with age in the EM (n = 31), END (n = 21) and ECT (n = 22). b,c, Correlation of percentage of ILC1s (b) and ILC3s (c) in each tissue (EM, n = 31; END, n = 20; ECT, n = 22). d, Comparison of the percentage of ILCs in mononuclear cells between donors with and without prior pregnancies (EM, n = 30; END, n = 21; ETC, n = 21). e, Correlation of the percentage of ILCs in mononuclear cells in each tissue with time since last pregnancy. f, Correlation of percentage of CD161+ ILCs in each tissue with time since last pregnancy (pregnant: EM, n = 22; END, n = 13; ECT, n = 15; never-pregnant: EM, n = 6; END, n = 6; ECT, n = 6). g, Correlation of percentage of CD161+ ILCs in each tissue age (EM, n = 23; END, n = 18; ECT, n = 15). h, Comparison of CD161 expression between never-pregnant and previously pregnant patients (1+ pregnancies: EM, n = 23; END, n = 15; ECT, n = 15; no pregnancies: EM, n = 6; END, n = 6; ECT, n = 6). Each dot represents a different individual. Horizontal lines represent the median ± IQR. Spearman rank test for all correlations; non-parametric Mann–Whitney U-test for comparisons of two groups. All tests were two-sided.
The CD161 ligand (LLT1) is highly expressed on trophoblastic cells in the placenta33, so we investigated whether CD161 expression on ILCs was modulated by reproductive parameters. CD161 expression on endometrial ILCs negatively correlated with time after pregnancy, suggesting that as time passes from pregnancy, genital ILCs lose expression of CD161 (Fig. 5f). Interestingly, CD161 expression was not modified with age (Fig. 5g), suggesting that regulation of CD161 expression over time is associated with pregnancy only. To further explore this possibility, we compared CD161 expression in previously pregnant versus never-pregnant donors. We found that ILCs from previously pregnant donors in the ECT expressed higher levels of CD161 (Fig. 5h). These results show that genital ILCs are modified throughout the lifespan and with reproductive function in a compartmentalized manner in the FGT.
ILCs rapidly respond to HIV stimulation with transcriptional changes
To define the antiviral potential of genital ILCs, we performed CITE-seq on enriched genital mixed-cell suspensions before and after in vitro HIV stimulation for 30 min. This approach allowed us to determine the single-cell whole transcriptome profile along with surface protein expression concurrently immediately following HIV exposure and bypass fluorescence-activated cell-sorting, a prolonged process that can modify the phenotype of immune cells sensitive to their tissue microenvironment34.
To identify ILCs, we selected immune cells (CD45+; encoded by PTPRC) and filtered out cells positive for lineage markers based on surface protein expression (CD3, CD11b, CD11c, CD14, CD15, CD16 and CD19) (Fig. 6a). Within this population, we selected three clusters of interest that expressed IL7R for further analysis (clusters 3, 4 and 5) (Fig. 6b). We validated our selection by confirming expression of previously described ILC genes, such as KLRB1, ZNF683 and ITGAE15 (Fig. 6c), and exclusion of genes associated with NK cell transcriptional signatures, such as EOMES, NCAM1 and NCR1 (ref. 35), which were enriched in IL7R-low clusters (clusters 0, 1, 2, 6; Fig. 6d). This indicated that clusters 3, 4 and 5 are enriched for helper ILCs, and we proceeded to further characterize these ILC-enriched clusters.

a, Representative uniform manifold approximation and projection (UMAP) visualization of lineage-negative genital cells. b, UMAP plot of IL7R expression in lineage-negative genital cells from the EM, END and ECT. c,d, Gene expression of common human ILC (c) and NK-cell-associated genes (d). e–h, Gene expression for ILC1 (e), intraepithelial ILC1 (f), ILC3 (g) and LTI-like cells (h). i, Volcano plot of differentially expressed genes between control and HIV. Paired comparisons were done with the Wilcoxon non-parametric test (two-sided). Red indicates P ≤ 0.05. j,k, Transcripts significantly upregulated (j) and downregulated (k) in response to HIV in genital ILCs. Red denotes HIV-stimulated; blue represents control. All have adjusted P ≤ 0.05. l, Gene Ontology terms from upregulated genes (P ≤ 0.05) with HIV stimulation. Fisher’s exact test with false discovery rate (FDR) correction. FDR is shown by color; bubble size is the number of genes. n = 3 tissues (EM, END, ECT).
We determined that clusters 3, 4 and 5 contained both ILC1 and ILC3 signatures (Fig. 6e–h). ILC1-associated genes such as ITGA1, ZBTB16, IKZF3 and TBX21 (refs. 14,15) were expressed across the uniform manifold approximation and project plot of genital ILCs (Fig. 6e). We also identified signature genes of intraepithelial ILC1s, such as PRDM1 and CD160 (Fig. 6f), previously described in the gut15. ILC3 signature genes were also detected throughout (KIT, AHR, CCR6 and LTB)14,15 (Fig. 6g). Additionally, genital ILCs expressed lymphoic tissue inducer (LTi)-associated genes such as GPR183 and NRP1 (refs. 36,37) (Fig. 6h).
To determine whether HIV exposure induces transcriptional changes in genital ILCs, we performed differential gene expression analysis between control and HIV-stimulated conditions in our ILC clusters. We identified genes upregulated and downregulated within 30 min following HIV exposure (Fig. 6i). Genital ILCs upregulated expression of PRIM1 and CXCL8, which are important for cell proliferation and chemoattraction of myeloid cells, respectively. We also detected significant upregulation of PLEK (pleckstrin) (Fig. 6j), encoding a substrate for PKC in leukocytes, and an important intermediate in the secretion and activation pathways of proinflammatory cytokines like TNF and IL-1β38. Consistently, genital ILCs significantly upregulated IL1B and TNF, as well as IL1R2, encoding the IL-1 decoy receptor (Fig. 6j). Additionally, ILCs upregulated BCL2A1, encoding an anti-apoptotic protein, and CD52, encoding a surface glycoprotein involved in lymphocyte migration and activation. ROMO1, encoding a protein that increases reactive oxygen species (ROS), was also upregulated, suggesting ILCs respond to HIV through the increase in ROS (Fig. 6j).
We also identified genes downregulated with HIV stimulation (Fig. 6k). Genital ILCs downregulated expression of CYGB and RHOBTB3, both involved in limiting ROS production (Fig. 6k). Genes involved in preventing proliferation (UBA7 and CNOT3) and cell death (NINJ1) were also downregulated. DGAT1 expression, involved in the formation of lipid droplets responsible for creating an immunosuppressive environment, was also downregulated (Fig. 6k).
To further understand the overall genital ILC response to HIV, we performed Gene Ontology analysis of the statistically significant genes (P ≤ 0.05) upregulated after stimulation and expressed in at least 10% of cells. Analysis of the upregulated genes, such as IL7R, TNF, NLRP3, CD226 and SLC11A1, contributed to a term associated with regulation of the adaptive immune response through somatic recombination of immune receptors. We detected many pathways associated with innate immune processes. Genes like NCKAP1L, IL1B, CXCL8, PDE4B and CSF3R contributed to biological processes related to neutrophil chemotaxis (Fig. 6l). HCK, IL1B, PDE4B, TNF and TICAM1 were involved in the term cellular response to lipopolysaccharide, suggesting potential activation of TLR4 by HIV39. We also detected terms associated with cell surface signaling, indicating that ILCs are undergoing an immediate response to the virus. Additionally, multiple pathways associated with ROS production were upregulated in response to HIV (Fig. 6l).
Overall, in response to HIV stimulation, genital ILCs triggered transcriptional signatures involved in inflammatory responses (cytokines and ROS), innate immune responses and cell proliferation.
ILCs respond to HIV through cytokine secretion and surface receptor modulation
Given that our data demonstrate that ILCs transcriptionally respond to HIV, we wanted to characterize the interactions between HIV and ILCs and the elicited responses at the protein level. First, we investigated HIV receptors. ILCs are not considered HIV-target cells, and whether they express HIV receptors is controversial23,40. Our flow cytometry analysis (Fig. 2c) demonstrated the presence of CD4+ ILCs in the FGT. Therefore, we further investigated HIV receptor expression in genital ILCs in a larger cohort. ILCs expressed CD4 at low levels compared to CD4+ T cells from the same tissue (Fig. 7a). CD4 expression was enriched on ILCs from the END and ECT compared to the EM (Fig. 7a,b) and was preferentially expressed on ILC1s (Fig. 7c). CD4+ ILC1s were primarily CD161 low or negative (Fig. 7d). To verify our flow cytometry results with untouched cells, we investigated CD4 expression through ChipCytometry and identified CD4+ ILC1s in the ECT (Fig. 7e). Evaluation of classical co-receptors showed that CCR5 was expressed by a portion of ILCs, with no tissue differences, but enriched on ILC1s (Extended Data Fig. 5a). By contrast, CXCR4 was broadly expressed in all ILCs and enriched in the ECT compared to the EM (Extended Data Fig. 5b). CD4/CCR5 co-expression is critical for mucosal HIV acquisition41. We detected a small percentage of ILCs co-expressing both receptors, primarily in END and ECT (Fig. 7f), suggesting potential viral entry through classical HIV receptors in a small subset of genital ILCs.

a, Representative contour plot of CD4 expression in genital T cells, EM, END and ECT ILCs. b, Percentage of ILCs expressing CD4 for each tissue type across all patients (n = 60 tissues: EM, n = 24; END, n = 19; ECT, n = 19). c, Percentage of CD4+ ILC1s and ILC3s in each tissue (n = 60 tissues: EM, n = 24; END, n = 18; ECT, n = 19). d, Comparison of CD161 MFI between CD4+ and CD4− ILCs (n = 60 tissues). e, Representative image of a CD4+ ILC1 in the ECT. f, Representative contour plot of CD4 and CCR5 co-expression on ILCs (left) and quantification (right) (EM, n = 24; END, n = 24; ECT, n = 19). g, Histogram of IFNγ (top) and CD107a (bottom) expression in CD161+ and CD161− ILC1s. Quantification of intracellular IFNγ and CD107a in control and HIV-stimulated ILC1s (n = 9). The connecting line represents one patient. h, Contour plot showing CD4 and CD107a expression in control ILC1s and stimulated with HIV and gp120. Quantification of CD4+ ILC1s with and without HIV stimulation (right) (n = 10 tissues). i, Percentage of CD161− ILC1s that were CD4+ (left), IFNγ+ (middle) and CD107a+ before and after gp120 stimulation (n = 9 tissues) j, Histogram of IL-22 (top) and CD107a (bottom) expression in CD161+ and CD161− ILC3s. Quantification of intracellular IL-22 and CD107a in control and HIV-stimulated ILC3s (right). The connecting line represents one patient (n = 9). k, Contour plot representing CD161 and NKp44 expression on endometrial (top left) and ectocervical (bottom left) ILC3s stimulated with and without HIV. Percentage of CD161hi (top right) (n = 12 tissues) and NKp44+ (n = 6 tissues) (bottom right) ILC3s with HIV stimulation. l, Percentage of IL-22+ (top) and CD107a+ (bottom) CD161+ ILC3s with and without gp120 stimulation (n = 8 tissues). Each dot represents a biological replicate. Horizontal lines represent the median ± IQR. Statistical analysis was performed using the non-parametric Kruskal–Wallis test (two-sided) with Dunn’s post hoc test correction for multiple comparisons. Paired comparisons were done with the Wilcoxon non-parametric test (two-sided).
Next, we investigated initial ILC responses to HIV challenge (30 min) at the protein level. This time was chosen to reveal immediate ILC responses to HIV before viral infection or replication and determine whether genital ILCs can use their pre-formed cytokines as a first-line defense in antiviral responses.
After HIV challenge, we detected a reduction in intracellular IFNγ content together with increased degranulation (CD107a expression) in ILC1s (Fig. 7g). Interestingly, these changes were specific to CD161− ILC1s, the subset with high intracellular cytokine content at homeostasis (Fig. 3b), but we detected no response in CD161+ ILC1s (Fig. 7g). IFNγ intracellular expression in CD161⁻ ILC1s following HIV stimulation decreased by 83.15% relative to control (Extended Data Fig. 5c). Additionally, granularity of CD161⁻ ILC1s, but not in CD161+ ILC1s, significantly decreased (measuring side scatter area; Extended Data Fig. 5d), suggesting granular release. Furthermore, genital ILC1s downregulated CD4 expression after HIV stimulation (Fig. 7h). As a control, genital T cells in the same sample did not downregulate CD4 upon 30 min of HIV stimulation (Extended Data Fig. 5e). To determine the mechanism for viral recognition, we performed stimulations with a CCR5-tropic gp120, the HIV glycoprotein that binds to CD4 and CCR5 (ref. 42). Following gp120 stimulation, we observed a significant increase in degranulation (CD107a) in genital CD161− ILC1s, but only a non-significant trend for decreased intracellular IFNγ content and no CD4 downregulation (Fig. 7i), suggesting that engagement of HIV receptors on ILC1s is only partially responsible for the observed effects.
Next, we evaluated ILC3 responses to HIV. Following HIV stimulation, CD161+ ILC3s decreased intracellular IL-22 content, but not CD161− ILC3s (Fig. 7j). CD161⁺ ILC3s exhibited about a 49.4% decrease in intracellular IL-22 following HIV stimulation relative to control (Extended Data Fig. 5c). In contrast to ILC1s, ILC3s did not increase CD107a expression upon HIV stimulation (Fig. 7j), but granularity (side scatter area) was reduced in CD161+ ILC3s (Extended Data Fig. 5d), suggesting that IL-22 release is independent of CD107a. Additionally, we detected CD161 expression downregulation concurrently with NKp44 upregulation (Fig. 7k), suggesting a shift towards an activated phenotype43. Importantly, stimulation with gp120 did not elicit any IL-22 or CD107a responses on ILC3s (Fig. 7l), which lack CD4 expression (Fig. 7a). This lack of response suggests that recognition and response to HIV by ILC3s is not mediated through classical HIV receptors. Finally, we did not detect any changes in intracellular cytokines (IFNγ, IL-22) or degranulation (CD107a) in T cells or NK cells from the same samples within 30 min (Extended Data Fig. 5f), further emphasizing the significance of the prompt response of ILCs to HIV stimulation.
Discussion
We investigated the role of ILCs in human mucosal immunity within the FGT. We defined multiple genital ILC subpopulations and unraveled previously unknown anatomical compartmentalization of subsets, spatial distribution and function. Our study demonstrates constitutive production of cytokines and degranulation in a subset-specific and tissue-specific manner at homeostasis and demonstrates the ability of genital ILCs to respond to HIV challenge immediately upon exposure throughout the FGT. These findings uncover critical roles for human genital ILCs in controlling mucosal immunity at homeostasis and initiating antiviral responses following HIV exposure.
The genital mucosa is unique and different from other mucosal surfaces owing to the tight, cyclical hormonal regulation of immune cells to optimize conditions for reproduction7. Despite the indication from a limited number of studies that ILCs may be important for pregnancy, ILCs in the female genital mucosa (human and murine) remain vastly uncharacterized. In humans, helper ILCs have only been partly characterized in the EM or during pregnancy21,22, but their role in mucosal protection during homeostasis and in response to infection throughout the FGT is completely unknown. Here we identified two genital ILC1 subpopulations (CD103+ and CD103−) and three ILC3 subpopulations (CCR6+CD103+, CCR6+CD103− and CCR6−) with tissue-dependent adaptations. Although CD103+ ILC1s may represent intraepithelial ILC1s as described previously in the gut44, genital ILC3s also express high levels of CD103 throughout. These data highlight the importance of studying each human mucosal site independently, as even within the FGT, ILCs possess tissue-specific phenotypes.
In our study, we uncovered that ILC1s and ILC3s constitutively produce IFNγ and IL-22, respectively, in the absence of stimulation. This production of cytokines seems to be enhanced in the FGT compared to other mucosal surfaces, as other studies reporting cytokine production by ILCs from lung12, gut13 and tonsil18 required in vitro stimulation to detect cytokines. Given that IFNγ and IL-22 are important for antiviral responses and epithelial barrier maintenance, our findings suggest that genital ILCs are preloaded with cytokines critical to provide an antiviral tissue state before pathogen exposure and represent a rapid source of cytokines to initiate antiviral responses. We speculate that this phenotype may reflect a specialized homeostatic state shaped by the unique tissue environment and immunological demands of the FGT, which must balance tolerance for sperm and potential embryo implantation with defense against pathogens. This environment is subject to repeated microbial, inflammatory and hormonal fluctuations across the menstrual cycle, including cyclical changes in estrogen and progesterone that are known to modulate immunity and barrier integrity5,7. These factors may help maintain a partially activated ILC state that supports reproductive capacity while protecting tissue homeostasis. Future studies should examine how different hormonal, reproductive and inflammatory factors influence genital ILCs to determine the underlying mechanisms responsible for this ‘primed’ state in the FGT.
Moreover, we identified that CD161 expression was associated with high cytokine content in an opposite fashion between ILC1s and ILC3s. Although the role of CD161 in ILCs remains unclear, prior studies on NK cells indicate that engagement of CD161 with its ligand results in inhibited cytotoxicity and IFNγ secretion45. This would fit with our finding that CD161− ILC1s have increased production of IFNγ, but how CD161 expression influences ILC3 production of IL-22 deserves further investigation. Importantly, our findings provide a targetable molecule (CD161) to investigate the regulation of cytokine production by genital ILCs.
Another finding in our study is the identification of a unique LTi-like endometrial ILC3 subset (NKp44+CCR6+CD103−) in adult human EM that constitutively degranulates at homeostasis and is located within endometrial lymphoid structures, interacting closely with T cells and B cells. Interestingly, contrary to our findings, prior studies in decidua reported that NKp44− ILC3s were more LTi-like than NKp44+ ILC3s21. Unlike other mucosal surfaces, the female genital mucosa is not a classical mucosal inductive site, and the majority of T cells present are tissue-resident memory phenotype28,29,46. A previous porcine study has suggested that engagement of the natural cytotoxicity receptor NKp44 can lead to degranulation47. Location within lymphoid aggregates paired with constitutive degranulation selectively in the EM suggests that endometrial NKp44+ ILC3s are degranulating within lymphoid aggregates after engagement of ligands on T cells and B cells, potentially to control their function. It remains to be determined what NKp44+ ILC3s release through the CD107a-mediated degranulation pathway. Prior studies described lymphoid aggregate structures in the EM composed of B cells and memory T cells that cyclically develop during the menstrual cycle48. Whether sex hormones control NKp44+ ILC3s to drive the formation and disassembly of lymphoid aggregates warrants further investigation. The presence of NKp44+ ILC3s in these lymphoid aggregates, along with their degranulation capabilities, highlights their potential role in orchestrating immune responses outside of conventional cytokine secretion.
Interestingly, our analysis revealed changes in ILC diversity with age and prior pregnancies. These data support prior studies pointing to the involvement of ILCs in reproduction21,22, but our study adds a level of complexity by uncovering dynamic changes in the human endometrial immune environment between pregnancies. Age and reproductive history represent a source of inter-individual variability that may influence the functional properties of genital ILCs under homeostasis and in response to pathogens, with potential consequences for innate mucosal protection and susceptibility to infections. Future studies need to determine whether sex hormones or specific tissue factors induced during pregnancy influence ILC plasticity.
Given that the initial immune responses triggered immediately after mucosal HIV exposure cannot be dissected in vivo, we investigated these responses using an in vitro model of mucosal HIV acquisition. Although ILCs have been studied in chronic HIV infection (in blood, gut and tonsils)18,23,24, their potential involvement during initial mucosal exposure has not been investigated. Our study provides evidence that ILCs respond to HIV immediately after stimulation and represent a rapid source of effector cytokines in the FGT, a major portal of entry for HIV acquisition. Importantly, CD161 expression was a predictor of cytokine secretion by ILCs preloaded at homeostasis. IFNγ could have antiviral actions at multiple levels, including downregulation of CD4 to inhibit HIV entry into cells49. Paired with our imaging evidence that ILC1s are tightly associated with T cells in the genital mucosa, IFNγ production by ILC1s may protect T cells from infection. However, we also observed rapid CD4 downregulation on ILC1s following HIV stimulation, which could suggest viral internalization and susceptibility to infection. Whether ILC1s are targets for viral infection and replication remains controversial23,40 and deserves further investigation, as they may represent previously unrecognized targets for HIV acquisition. Additionally, although ILC3s did not express classical HIV receptors, we detected antiviral responses with a rapid switch from a homeostatic CD161+ phenotype to an activated NKp44+ phenotype. Interestingly, in chronically simian immunodeficiency virus-infected animals, NKp44+ ILCs negatively correlated with viral load, suggesting a protective effect of NCR+ ILCs50,51.
As our HIV stimulation assays were performed in the context of mixed-cell suspensions to mimic the mucosal environment, it remains undetermined whether genital ILCs directly recognize and respond to HIV or indirectly respond to inflammatory signals and secreted molecules from surrounding cells. Our attempts to purify genital ILCs resulted in low cell numbers, low viability, impure populations and artificial activation, as determined by increased CD107a expression and decreased intracellular cytokine content after the selection process. These technical issues represent a limitation of our study that prevented us from directly addressing this question and need to be overcome in future studies to obtain pure populations that accurately represent the functional state of ILCs in genital tissues. However, regardless of whether the response is direct or indirect, our study demonstrates that ILCs present in the genital mucosa are rapid responders following mucosal HIV exposure. Furthermore, although we speculate that ILC-mediated IFNγ and IL-22 responses could have a protective role, the biological role of this response for mucosal protection against HIV in vivo remains to be determined.
Overall, we demonstrate unique plasticity, location and functional roles for ILCs in the FGT at homeostasis, along with their ability to immediately respond to HIV challenge. These insights and our detailed phenotypic analysis pave the way for future research into the role of ILCs in human mucosal immunity and their potential involvement in HIV pathogenesis.
Methods
Study subjects
Studies were performed with Tufts University and Wayne State University Institutional Review Board approval. HIV-negative women undergoing hysterectomies for benign conditions at Tufts Medical Center or Detroit Medical Center gave written informed consent before surgery. Approval to use tissues was obtained from the Committee for the Protection of Human Subjects. Hysterectomy samples from three patients were obtained through the National Disease Research Interchange. Tissue samples distal from the sites of pathology and without pathological lesions were selected by pathologists. De-identified information about age, surgery indication, menstrual or menopausal status and reproductive history was collected. For patients obtained through the National Disease Research Interchange, only information about age was available. General characteristics of the patients are shown in Supplementary Table 1.
Tissue processing
Tissues obtained from hysterectomies included EM, END and ECT (n = 110 tissues from n = 46 patients) and were transferred to the laboratory immediately after surgery. Tissues were processed to obtain stromal mixed-cell suspensions with optimized protocols to preserve tissue intrinsic features of resident cells, as previously described4,8. In brief, enzymatic digestion was performed in a gentleMACS Dissociator (Miltenyi Biotec) using the Tumor Dissociation Kit, Human (Miltenyi Biotec) and 0.01% DNase (Worthington Biochemical). After digestion, the tissues were filtered through 100 μm, 70 μm and 30 μm filters (MACS SmartStrainers; Miltenyi Biotec) to separate epithelial cells from stromal cells. Cell suspensions then underwent red blood cell removal using human CD235a (Glycophorin A) MicroBeads (Miltenyi Biotec). Mixed-cell suspensions were used for high-dimensional spectral flow cytometry analysis.
Flow cytometry analysis
Mixed-cell suspensions were stained for surface markers with combinations of anti-human antibodies shown in Supplementary Table 2. Dead cells were excluded with the LIVE/DEAD Fixable Blue Dead Cell Stain Kit (Invitrogen). Following surface marker staining, intracellular staining was performed by washing, fixing and permeabilizing cells (20 min), following instructions provided in the Cytofix/Cytoperm Plus kit (BD Biosciences), and then staining for intracellular cytokines for 30 min. For samples that underwent transcription factor staining, after surface staining, cells were fixed and permeabilized using the eBioscience Foxp3/Transcription Factor Staining Kit and stained for transcription factors for 30 min. Analysis was performed on a spectral flow cytometer, the five-laser Aurora cytometer (Cytek Biosciences) or the three-laser Northern Lights (Cytek Biosciences). Data were analyzed with OMIQ software (www.omiq.ai). Expression of surface markers is shown as a percentage of positive cells and the mean fluorescence intensity. The fluorescence-minus-one strategy was used to establish appropriate gates. Unsupervised clustering analysis was performed using OMIQ software (www.omiq.ai). Data from three patients with matching FGT tissues (EM, END and ECT) were manually gated as shown in Fig. 1a to identify total ILCs (lineage-negative CD127+). Then, opt-SNE52, a dimensionality reduction method, was applied for visualization, followed by FlowSOM53, a clustering algorithm to define genital ILC clusters based on marker expression.
Flow cytometry-based cytokine secretion assay
Secretion of IL-22 was determined using the flow-cytometry-based IL-22 Secretion Assay–Detection Kit (PE) (Miltenyi). The kit was used according to the manufacturer’s instructions with modifications. Mixed-cell suspensions were generated and incubated on ice with the IL-22 catch reagent. After 5 min, the CD45 Spark NIR 685 antibody was added to the mixed-cell suspension, and cells were diluted in warm media as recommended. Cells were then stimulated with the Cell Activation Cocktail without Brefeldin A (a pre-mixed cocktail with PMA and ionomycin) (BioLegend) according to the manufacturer’s instructions. CD107a antibody was added, and the cells were incubated for 1 h at 37 °C with slow rotation as recommended. After 1 h, cells were washed according to the manufacturer’s instructions and incubated with the IL-22 detection antibody for 10 min as recommended. After 10 min, cells were labeled with the LIVE/DEAD Fixable Blue Dead Cell Stain Kit (Invitrogen) and incubated with antibodies for cell surface markers to detect ILCs as detailed in the ‘Flow cytometry analysis’ section.
ChipCytometry
The CellScape Precise Spatial Multiplexing platform was used for automated tissue staining and imaging (Canopy Biosciences, A Bruker Company). Tissues were fresh-frozen in OCT and sectioned before staining. In brief, tissue sections were loaded into CellScape Chips following the manufacturer’s instructions. Tissues were stained with two to three fluorescently labeled antibodies at a time, followed by high-dynamic-range image acquisition of each fluorophore at a resolution of 182 nm per pixel. Fluorescent signal from all fluorophores was quenched through filtered photobleaching. Tissues were then washed with wash buffer and subjected to additional rounds of staining, imaging and photobleaching until all markers were completed. The antibodies used for staining are shown in Supplementary Table 3. Quantification of the ILC3 subset location was done by selecting regions of interest in which ILC3s were present, and either contained lymphoid aggregates or did not. For ILC1 subset location quantification, regions of interest were categorized as intraepithelial, subepithelial or deeper tissue, and the presence of different ILC1 subsets was compared between these regions. The number of cells per area was counted. Areas with no detected ILCs were assigned an arbitrary value of 0.1 (instead of 0) to allow representation on a log scale.
HIV viral growth
HIV-1-BaL (R5) isolates were acquired from the AIDS Research and Reference Reagent Program at the Division of AIDS, National Institute of Allergy and Infectious Diseases, National Institutes of Health, with contributions from S. Gartner, M. Popovic and R. Gallo54, and were propagated in activated peripheral blood mononuclear cells as previously described55.
HIV and gp120 stimulation
For in vitro HIV-stimulation experiments, mixed-cell suspensions were stimulated with HIV-1 BaL for 30 min at a multiplicity of infection of 0.5, after which residual virus was washed away. After washing, cells were stained for surface markers as indicated, followed by fixation and permeabilization (as described in the ‘Flow cytometry analysis’ section) and intracellular cytokine staining for 30 min. Data were acquired with the five-laser Aurora (Cytek Biosciences) or the three-laser Northern Lights (Cytek Biosciences). Stimulation with Recombinant BaL gp120 (National Institutes of Health HIV Reagent Program, cat. no. 4961) at 1 µg ml−1 was performed using the same methods as described above.
Sample preparation for multi-omic single-cell RNA sequencing with HIV stimulation
Single-cell transcriptome and surface antibody sequencing was performed using the BD Rhapsody platform. Mixed single-cell suspensions from hysterectomy samples obtained from three genital tissues (EM, END and ECT) were enriched for innate immune cells by magnetic bead removal of CD3+, CD19+, CD235a+ red blood cells and fibroblasts as previously described8. Cells were incubated with HIV-BaL (multiplicity of infection, 0.5) in XVIVO-15 or media alone for 30 min. Cells were washed to remove free virus and then incubated with oligo-conjugated antibodies (Supplementary Table 4) and barcoded sample tags. Cells were then washed twice in MACS buffer to remove unbound antibodies and sample tags. Cells were counted and loaded onto separate BD Rhapsody cartridges according to condition (HIV-stimulated or unstimulated controls). Next, polyA tail capture beads with unique molecular identifier barcodes were added to the microwells, followed by a lysis step to extract cellular RNA. Sequencing libraries were then created according to the manufacturer’s instructions.
Single-cell CITE-seq analysis
FASTQ files with unaligned reads were produced using the NovaSeq6000 instrument. Gene counts were obtained by aligning and annotating the reads to the human genome. Downstream analyses were accomplished using the Seurat R software package (v.5.1; http://satijalab.org/seurat). After filtering out cells with a high mitochondrial content and low number of features, we performed scaling and normalization using the SCTransform algorithm. Principal component analysis was used to reduce the dimensionality of the data. Uniform manifold approximation and projection was used for visualization. To identify genital ILCs, we filtered out cells with elevated expression of CD3, CD19, CD11b, CD11c, CD16, CD14 and CD15 surface proteins. Next, we identified ILC-enriched cell clusters through IL7R expression. Differentially expressed genes were identified from control and HIV-stimulated cells using the ‘FindMarkers’ function, considering genes that were found in at least 10% of the cells for at least one condition. Gene Ontology biological processes were produced with the Gene Ontology resource (https://geneontology.org) using a gene list of upregulated genes (P ≤ 0.05) with HIV stimulation, and pathways with a false discovery rate of ≤0.05 were chosen for visualization.
Statistics
Data analysis was performed using the GraphPad Prism 9.0 software. Comparison of two groups was performed with the non-parametric Mann–Whitney U-test or Wilcoxon paired test. Comparison of three or more groups was performed by applying the non-parametric Kruskal–Wallis test followed by Dunn’s post hoc test to correct for multiple comparisons. A two-sided P value of <0.05 was considered statistically significant; exact P values are provided on the figures.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
CITE-seq data have been deposited in the NCBI Gene Expression Omnibus repository under accession number GSE279775. Source data are provided with this paper.
Code availability
No custom code was generated in this study.
References
Women and girls and HIV (2018). UNAIDS https://www.unaids.org/en/resources/documents/2018/women_girls_hiv (2018).
Boily, M. C. et al. Heterosexual risk of HIV-1 infection per sexual act: systematic review and meta-analysis of observational studies. Lancet Infect. Dis. 9, 118–129 (2009).
Barr, F. D., Ochsenbauer, C., Wira, C. R. & Rodriguez-Garcia, M. Neutrophil extracellular traps prevent HIV infection in the female genital tract. Mucosal Immunol. 11, 1420–1428 (2018).
Moreno de Lara, L. et al. Aging dysregulates neutrophil extracellular trap formation in response to HIV in blood and genital tissues. Front. Immunol. 14, 1256182 (2023).
Rodriguez-Garcia, M., Connors, K. & Ghosh, M. HIV pathogenesis in the human female reproductive tract. Curr. HIV/AIDS Rep. 18, 139–156 (2021).
Rodriguez-Garcia, M. et al. Dendritic cells from the human female reproductive tract rapidly capture and respond to HIV. Mucosal Immunol. 10, 531–544 (2017).
Wira, C. R., Rodriguez-Garcia, M. & Patel, M. V. The role of sex hormones in immune protection of the female reproductive tract. Nat. Rev. Immunol. 15, 217–230 (2015).
Parthasarathy, S. et al. Human genital dendritic cell heterogeneity confers differential rapid response to HIV-1 exposure. Front. Immunol. 15, 1472656 (2024).
Vivier, E. et al. Innate lymphoid cells: 10 years on. Cell 174, 1054–1066 (2018).
Panda, S. K. & Colonna, M. Innate lymphoid cells in mucosal immunity. Front. Immunol. 10, 861 (2019).
Piersma, S. J. Tissue-specific features of innate lymphoid cells in antiviral defense. Cell Mol. Immunol. 21, 1036–1050 (2024).
De Grove, K. C. et al. Characterization and quantification of innate lymphoid cell subsets in human lung. PLoS ONE 11, e0145961 (2016).
Saez, A. et al. Innate lymphoid cells in intestinal homeostasis and inflammatory bowel disease. Int. J. Mol. Sci. 22, 7618 (2021).
Mazzurana, L. et al. Tissue-specific transcriptional imprinting and heterogeneity in human innate lymphoid cells revealed by full-length single-cell RNA-sequencing. Cell Res. 31, 554–568 (2021).
Jaeger, N. et al. Diversity of group 1 innate lymphoid cells in human tissues. Nat. Immunol. 25, 1460–1473 (2024).
Yudanin, N. A. et al. Spatial and temporal mapping of human innate lymphoid cells reveals elements of tissue specificity. Immunity 50, 505–519.e4 (2019).
Pascual-Reguant, A. et al. Multiplexed histology analyses for the phenotypic and spatial characterization of human innate lymphoid cells. Nat. Commun. 12, 1737 (2021).
Singh, A. et al. Innate lymphoid cell activation and sustained depletion in blood and tissue of children infected with HIV from birth despite antiretroviral therapy. Cell Rep. 32, 108153 (2020).
Cella, M. et al. Subsets of ILC3–ILC1-like cells generate a diversity spectrum of innate lymphoid cells in human mucosal tissues. Nat. Immunol. 20, 980–991 (2019).
Bruggen, M. C. et al. In situ mapping of innate lymphoid cells in human skin: evidence for remarkable differences between normal and inflamed skin. J. Invest. Dermatol. 136, 2396–2405 (2016).
Vacca, P. et al. Identification of diverse innate lymphoid cells in human decidua. Mucosal Immunol. 8, 254–264 (2015).
Doisne, J. M. et al. Composition, development, and function of uterine innate lymphoid cells. J. Immunol. 195, 3937–3945 (2015).
Kloverpris, H. N. et al. Innate lymphoid cells are depleted irreversibly during acute HIV-1 infection in the absence of viral suppression. Immunity 44, 391–405 (2016).
Kramer, B. et al. Compartment-specific distribution of human intestinal innate lymphoid cells is altered in HIV patients under effective therapy. PLoS Pathog. 13, e1006373 (2017).
Werner, A., Holmes, A., Moldovan, G. & Rodriguez-Garcia, M. Innate lymphoid cells in HIV pathogenesis and in the human female genital tract. Curr. Opin. HIV AIDS 20, 117–123 (2025).
Parthasarathy, S. et al. Aging modifies endometrial dendritic cell function and unconventional double negative T cells in the human genital mucosa. Immun. Ageing 20, 34 (2023).
Rodriguez-Garcia, M., Barr, F. D., Crist, S. G., Fahey, J. V. & Wira, C. R. Phenotype and susceptibility to HIV infection of CD4+ Th17 cells in the human female reproductive tract. Mucosal Immunol. 7, 1375–1385 (2014).
Rodriguez-Garcia, M., Fortier, J. M., Barr, F. D. & Wira, C. R. Aging impacts CD103+ CD8+ T cell presence and induction by dendritic cells in the genital tract. Aging Cell 17, e12733 (2018).
Rodriguez-Garcia, M., Shen, Z., Fortier, J. M. & Wira, C. R. Differential cytotoxic function of resident and non-resident CD8+ T cells in the human female reproductive tract before and after menopause. Front. Immunol. 11, 1096 (2020).
Metcalfe, D. D. et al. Biomarkers of the involvement of mast cells, basophils and eosinophils in asthma and allergic diseases. World Allergy Organ. J. 9, 7 (2016).
Ghosh, M. et al. CCL20/MIP3α is a novel anti-HIV-1 molecule of the human female reproductive tract. Am. J. Reprod. Immunol. 62, 60–71 (2009).
Kang, J. et al. Type 3 innate lymphoid cells are associated with a successful intestinal transplant. Am. J. Transpl. 21, 787–797 (2021).
Llibre, A. et al. Expression of lectin-like transcript-1 in human tissues. F1000Res 5, 2929 (2016).
Llufrio, E. M., Wang, L., Naser, F. J. & Patti, G. J. Sorting cells alters their redox state and cellular metabolome. Redox Biol. 16, 381–387 (2018).
Rebuffet, L. et al. High-dimensional single-cell analysis of human natural killer cell heterogeneity. Nat. Immunol. 25, 1474–1488 (2024).
Chu, C. et al. Anti-microbial functions of group 3 innate lymphoid cells in gut-associated lymphoid tissues are regulated by G-protein-coupled receptor 183. Cell Rep. 23, 3750–3758 (2018).
Shikhagaie, M. M. et al. Neuropilin-1 is expressed on lymphoid tissue residing LTi-like group 3 innate lymphoid cells and associated with ectopic lymphoid aggregates. Cell Rep. 18, 1761–1773 (2017).
Ding, Y. et al. Phosphorylation of pleckstrin increases proinflammatory cytokine secretion by mononuclear phagocytes in diabetes mellitus. J. Immunol. 179, 647–654 (2007).
Nazli, A. et al. HIV-1 gp120 induces TLR2- and TLR4-mediated innate immune activation in human female genital epithelium. J. Immunol. 191, 4246–4258 (2013).
Zhao, J. et al. Infection and depletion of CD4+ group-1 innate lymphoid cells by HIV-1 via type-I interferon pathway. PLoS Pathog. 14, e1006819 (2018).
Saba, E. et al. HIV-1 sexual transmission: early events of HIV-1 infection of human cervico-vaginal tissue in an optimized ex vivo model. Mucosal Immunol. 3, 280–290 (2010).
Yoon, V. et al. The GP120 molecule of HIV-1 and its interaction with T cells. Curr. Med. Chem. 17, 741–749 (2010).
Vitale, M. et al. NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non-major histocompatibility complex-restricted tumor cell lysis. J. Exp. Med. 187, 2065–2072 (1998).
Fuchs, A. et al. Intraepithelial type 1 innate lymphoid cells are a unique subset of IL-12- and IL-15-responsive IFN-γ-producing cells. Immunity 38, 769–781 (2013).
Aldemir, H. et al. Cutting edge: lectin-like transcript 1 is a ligand for the CD161 receptor. J. Immunol. 175, 7791–7795 (2005).
Shen, Z., Vom Steeg, L. G., Patel, M. V., Rodriguez-Garcia, M. & Wira, C. R. Impact of aging on the frequency, phenotype, and function of CD4+ T cells in the human female reproductive tract. Front. Immunol. 15, 1465124 (2024).
Mair, K. H. et al. The natural cytotoxicity receptor NKp44 (NCR2, CD336) is expressed on the majority of porcine NK cells ex vivo without stimulation. Front. Immunol. 13, 767530 (2022).
Yeaman, G. R., Collins, J. E., Fanger, M. W., Wira, C. R. & Lydyard, P. M. CD8+ T cells in human uterine endometrial lymphoid aggregates: evidence for accumulation of cells by trafficking. Immunology 102, 434–440 (2001).
Kang, S., Brown, H. M. & Hwang, S. Direct antiviral mechanisms of interferon-gamma. Immune Netw. 18, e33 (2018).
Rahman, M. A. et al. Differential effect of mucosal NKp44+ innate lymphoid cells and Δγ cells on simian immunodeficiency virus infection outcome in rhesus macaques. J. Immunol. 203, 2459–2471 (2019).
Rahman, M. A., Silva de Castro, I., Schifanella, L., Bissa, M. & Franchini, G. Vaccine induced mucosal and systemic memory NK/ILCs elicit decreased risk of SIV/SHIV acquisition. Front. Immunol. 15, 1441793 (2024).
Belkina, A. C. et al. Automated optimized parameters for T-distributed stochastic neighbor embedding improve visualization and analysis of large datasets. Nat. Commun. 10, 5415 (2019).
Van Gassen, S. et al. FlowSOM: using self-organizing maps for visualization and interpretation of cytometry data. Cytometry A 87, 636–645 (2015).
Gartner, S. et al. The role of mononuclear phagocytes in HTLV-III/LAV infection. Science 233, 215–219 (1986).
Rodriguez-Garcia, M. et al. Estradiol reduces susceptibility of CD4+ T cells and macrophages to HIV-infection. PLoS ONE 8, e62069 (2013).
Acknowledgements
We thank R. Wang for technical assistance. B. Malla, P. Josephs and A. Azimirad (Department of Gynecology and Obstetrics, Tufts Medical Center, Boston, MA) for their help with clinical coordination and patient recruitment and K. Murga (Biorepository at Tufts Medical Center) for tissue supply. This work was supported by the National Institutes of Health R01AG060801 (M.R.-G.), R21AI172065 (M.R.-G.), U01AG084766 (M.R.-G.) and Burroughs Wellcome Fund PATH award (1334236) (M.R.-G.). The David Thorley–Lawson Memorial Flow Cytometry Core Facility at Tufts University is supported by S10OD032201. The Microscopy, Imaging and Cytometry Resources Core at Wayne State is supported in part by National Institutes of Health Center grant P30 CA22453 to the Karmanos Cancer Institute and R50 CA251068-01 to K. Moin, Wayne State University.
Author information
Authors and Affiliations
Contributions
M.R.-G. and A.W. conceptualized the study. A.W., J.F. and M.R.-G. conducted the formal analyses. M.R.-G., A.W., L.M., A.H., S.P., A.B., F.C.S., J.F. and G.M. developed the methodology. A.W., L.M., A.H., S.P., A.B., F.C.S., G.M. and M.R.-G. performed the investigation. A.V., V.I., J.S. and R.J. procured resources. A.W., M.R.-G. and J.F. visualized the study. M.R.-G., A.V. and V.I. were responsible for project administration. M.R.-G. supervised the research and acquired funding. A.W. and M.R.-G. wrote the original draft of the paper. M.R.-G., A.W., J.F., G.M., L.M., A.B., F.C.S., S.P. and A.H. contributed to editing; all authors reviewed and approved the final version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Immunology thanks Tamara Tilburgs and the other, anonymous, reviewers for their contribution to the peer review of this work. Peer reviewer reports are available. Primary Handling Editor: P. Jauregui, in collaboration with the Nature Immunology team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Genital helper ILCs are distinct from NK cells.
(a) Contour plot of CD335 expression in ILCs (left) and NK cells (right). (b) Contour plot of CD94 expression vs CD127 expression in lineage negative cells. (c) Contour plot of CD56 expression in lineage negative cells (left) with CD94 expression overlayed in color (middle). Quantification of CD94+ Lineage negative CD56 + CD127- cells and ILCs (right) (n = 6 tissues). (d) Quantification of percent of ILCs expressing CD56 (left). Percent of CD56 + ILC1s and ILC3s across tissues (right). EM = 22, END = 15, ECT = 16. (e) Contour plots of CD203c expression in each tissue comparing total CD45+ (top) and genital ILCs (bottom). Quantification of CD203c expression in CD45+ and ILCs (right) (n = 9 tissues). (f) Contour plot of cells expressing CD34 in total CD45- (top) and genital ILCs (bottom) (n = 6). (g) Contour plots depicting percentage of CD123 expression in total CD45+ lineage- but including CD11c+ cells (left) and CD45+ lineage- CD11c- cells (middle). Quantification of CD123 expression (right) (n = 3 tissues) (h) Representative contour plot depicting CD294 and CD4 expression in CD3+ cells (left) and ILCs (right). Percent of CD294 + T-cells (CD3+) in different donors (n = 43 tissues). (i) CD127 MFI comparison between ILC1s and ILC3s in all tissues. (n = 56 tissues). (j) Representative contour plot of IL-5 (left), IL-17 (middle), and IL-13 (right) expression in genital ILCs. (k) Contour plot of Granulysin expression in genital ILCs (left) and NK cells (right) (n = 56 tissues). Each dot represents a biological replicate. Horizontal lines represent the median ± IQR. Statistical analysis was performed using the non-parametric Kruskal–Wallis test (two-sided) with Dunn’s post-test correction for multiple comparisons. Paired comparisons were done with the Wilcoxon non-parametric test. *P < 0.05; **P < 0.01; ****P < 0.0001.
Extended Data Fig. 2 Genital ILCs exhibit tissue- and patient-dependent protein expression differences.
(a) opt-SNE plots on concatenated total ILCs from 3 matched patients’ endometrium, endocervix, and ectocervix samples stratified by patient. (b) FlowSOM clustering analysis overlayed onto the opt-SNE plot with ten distinct clusters with matched ILC subset type. (c) Heatmap of the ten distinct clusters (rows) depicting the markers of interest with their relative expression values (red is high expression, blue is low). (d) MFI of CD107a (top), IL-22 (middle), and IFN-y (bottom) by cluster. Shapes indicate tissue type, while color indicates patient. (e) Dot plot with an FMO (left) used to show detection of CCR6 in CD45+ genital cells (right).
Extended Data Fig. 3 Functionality of ILC subsets.
(a) Contour plots depicting IFN-y expression in CD103+ (left) and CD103- (right) ILC1s. (b) Contour plots of IFN-y expression in CD161+ and CD161- ILC1s. (c) Contour plots of IL-22 expression in the three distinct ILC3 subsets, CCR6 + CD103-, CCR6 + CD103+, and CCR6-. (d) Contour plot of IL-22 expression in CD161+ and CD161- ILC3s. (e) Contour plot of CD107a expression in three ILC3 subsets and ILC1s. (f) Lack of correlation between percentage of IFN-y + ILC1s and IL-22 + ILC3s (top) (n = 32 tissues); endometrial CD107a + ILC3s and IL-22 + ILC3s (middle) (n = 11 tissues); and endometrial CD107a + ILC3s and IFN-y + ILC1s (bottom) (n = 11 tissues). Each dot represents one patient with both values. Spearman rank test (two-sided) for all correlations.
Extended Data Fig. 4 Quantification of genital ILCs with ChipCytometry.
(a) Representative fluorescent images depicting lineage markers and ILC markers to identify ILCs. (b) Representative example of selected regions of interest (ROI) for quantification of ILC3s in the endometrium according to location within lymphoid aggregates or outside lymphoid aggregates. Yellow depicts a representative ROI with a LA (lymphoid aggregate) and red depicts representative regions without lymphoid aggregates (non-LA). (c) Representative example of selected ROI in ectocervix for ILC1 quantification. Yellow depicts intraepithelial, red subepithelial, and white deeper tissue ROIs.
Extended Data Fig. 5 Genital ILCs respond to HIV.
(a) Contour plot of CCR5 expression in a fluorescent-minus-one (FMO), and genital ILCs (top). Percent of genital ILCs expressing CCR5 (bottom left), and percent of ILC1s and ILC3s expressing CCR5 (bottom right). EM (n = 17), END (n = 14), ECT (n = 15). (b) Contour plot of CXCR4 expression in T-cells and genital ILCs (top). Percent of genital ILCs expressing CXCR4 (bottom left), and percent of ILC1s and ILC3s expressing CXCR4 (bottom right). EM (n = 22), END (n = 17), ECT (n = 21). (c) Fold change in intracellular IFN-γ expression in CD161⁻ ILC1s (left) and IL-22 expression in CD161⁺ ILC3s (right) following HIV stimulation, normalized to their respective unstimulated controls (set to 1). Each line represents a matched individual tissue. Horizontal lines indicate the mean. Statistical significance was assessed using a one-sample Wilcoxon rank test. (d) Side scatter area comparison between control and HIV stimulated CD161+/- ILC1s and CD161+/- ILC3s. (n = 9 tissues) (e) CD4 and CD107a expression on CD3+ cells before and after HIV and gp120 stimulation. (f) Percent of IFN-y+ (left), IL-22+ (middle), and CD107a+ (right) NK cells (CD56 + CD127-) and T-cells (CD3+) with (white) and without (black) HIV stimulation. Wilcoxon test (two-sided). Each dot represents a patient (n = 9). Horizontal lines represent the median ± IQR. Statistical analysis was performed using the non-parametric Kruskal–Wallis test (two-sided) with Dunn’s post-test correction for multiple comparisons. *P < 0.05; **P < 0.01.
Supplementary information
Supplementary Tables (download XLSX )
Supplementary tables combined.
Source data
Source Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 5 (download XLSX )
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Werner, A., Moreno de Lara, L., Holmes, A. et al. Spatial and functional diversity of innate lymphoid cells in the human female genital tract may contribute to antiviral responses to HIV. Nat Immunol 26, 2030–2044 (2025). https://doi.org/10.1038/s41590-025-02285-6
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41590-025-02285-6
This article is cited by
-
Female ILCs on the spotlight
Nature Immunology (2025)
