
Abstract
Alzheimer’s disease causes progressive cognitive decline, yet some individuals remain resilient despite developing hallmark pathology. A subset of people with Down syndrome (DS), the most common genetic cause of Alzheimer’s disease, demonstrates such resilience. Given the elevated risk of hematopoietic mutations in DS, we hypothesize that certain variants may confer microglial resilience. Here, we introduce a myeloid DS-linked CSF2RB A455D mutation into human pluripotent stem cell-derived microglia from both donors with DS and healthy donors and study their function in 4–10-month-old chimeric mice. We find that this mutation suppresses type I interferon signaling in response to tau pathology, reducing inflammation while enhancing phagocytosis, thereby ameliorating microglial senescence. CSF2RB A455D-expressing microglia form a unique protective subpopulation and preserve neuronal functions. Importantly, they replace diseased wild-type microglia after tau exposure. These findings provide proof of concept that engineered human microglia can enhance resilience against tauopathy, opening avenues for microglial replacement therapies.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

Data availability
The RNA-seq, scRNA-seq and whole-genome sequencing datasets generated in this study have been deposited at the NCBI Gene Expression Omnibus (accession no GSE252238) and are publicly available as of the date of publication. Source data are provided with this paper.
Code availability
This paper does not report any original code.
References
Neuner, S. M. et al. Translational approaches to understanding resilience to Alzheimer’s disease. Trends Neurosci. 45, 369–383 (2022).
Zhang, M. et al. Resilience and resistance to the accumulation of amyloid plaques and neurofibrillary tangles in centenarians: an age-continuous perspective. Alzheimers Dement. 19, 2831–2841 (2023).
Rahimi, J. & Kovacs, G. G. Prevalence of mixed pathologies in the aging brain. Alzheimers Res. Ther. 6, 82 (2014).
Bouzid, H. et al. Clonal hematopoiesis is associated with protection from Alzheimer’s disease. Nat. Med. 29, 1662–1670 (2023).
Wilk, C. M. et al. Circulating senescent myeloid cells infiltrate the brain and cause neurodegeneration in histiocytic disorders. Immunity 56, 2790–2802 (2023).
Wiseman, F. K. et al. A genetic cause of Alzheimer disease: mechanistic insights from Down syndrome. Nat. Rev. Neurosci. 16, 564–574 (2015).
Lott, I. T. & Head, E. Dementia in Down syndrome: unique insights for Alzheimer disease research. Nat. Rev. Neurol. 15, 135–147 (2019).
Mann, D. M. Alzheimer’s disease and Down’s syndrome. Histopathology 13, 125–137 (1988).
Lemere, C. A. et al. Sequence of deposition of heterogeneous amyloid β-peptides and APO E in Down syndrome: implications for initial events in amyloid plaque formation. Neurobiol. Dis. 3, 16–32 (1996).
Leverenz, J. B. & Raskind, M. A. Early amyloid deposition in the medial temporal lobe of young Down syndrome patients: a regional quantitative analysis. Exp. Neurol. 150, 296–304 (1998).
Martini, A. C., Gross, T. J., Head, E. & Mapstone, M. Beyond amyloid: immune, cerebrovascular, and metabolic contributions to Alzheimer disease in people with Down syndrome. Neuron 110, 2063–2079 (2022).
Krinsky-McHale, S. J. et al. Successful aging in a 70-year-old man with down syndrome: a case study. Intellect. Dev. Disabil. 46, 215–228 (2008).
Osuna-Marco, M. P., López-Barahona, M., López-Ibor, B. & Tejera, Á. M. Ten reasons why people with Down syndrome are protected from the development of most solid tumors—a review. Front. Genet. 12, 749480 (2021).
Hitzler, J. K. & Zipursky, A. Origins of leukaemia in children with Down syndrome. Nat. Rev. Cancer 5, 11–20 (2005).
Mateos, M. K., Barbaric, D., Byatt, S.-A., Sutton, R. & Marshall, G. M. Down syndrome and leukemia: insights into leukemogenesis and translational targets. Transl. Pediatr. 4, 76–92 (2015).
Hasle, H., Clemmensen, I. H. & Mikkelsen, M. Risks of leukaemia and solid tumours in individuals with Down’s syndrome. Lancet 355, 165–169 (2000).
Roberts, I. et al. GATA1-mutant clones are frequent and often unsuspected in babies with Down syndrome: identification of a population at risk of leukemia. Blood 122, 3908–3917 (2013).
Wechsler, J. et al. Acquired mutations in GATA1 in the megakaryoblastic leukemia of Down syndrome. Nat. Genet. 32, 148–152 (2002).
Nikolaev, S. I. et al. Frequent cases of RAS-mutated Down syndrome acute lymphoblastic leukaemia lack JAK2 mutations. Nat. Commun. 5, 4654 (2014).
Yoshida, K. et al. The landscape of somatic mutations in Down syndrome-related myeloid disorders. Nat. Genet. 45, 1293–1299 (2013).
Gosselin, D. An environment-dependent transcriptional network specifies human microglia identity. Science 356, eaal3222 (2017).
Hansen, D. V., Hanson, J. E. & Sheng, M. Microglia in Alzheimer’s disease. J. Cell Biol. 217, 459–472 (2018).
Holtman, I. R., Skola, D. & Glass, C. K. Transcriptional control of microglia phenotypes in health and disease. J. Clin. Invest. 127, 3220–3229 (2017).
Labuhn, M. et al. Mechanisms of progression of myeloid preleukemia to transformed myeloid leukemia in children with Down syndrome. Cancer Cell 36, 123–138 (2019).
Broughton, S. E. et al. The βc receptor family—structural insights and their functional implications. Cytokine 74, 247–258 (2015).
Chitu, V. et al. Microglial homeostasis requires balanced CSF-1/CSF-2 receptor signaling. Cell Rep. 30, 3004–3019 (2020).
Boyd, T. D. et al. GM-CSF upregulated in rheumatoid arthritis reverses cognitive impairment and amyloidosis in Alzheimer mice. J. Alzheimers Dis. 21, 507–518 (2010).
Hamilton, J. A. GM-CSF in inflammation and autoimmunity. Trends Immunol. 23, 403–408 (2002).
Kao, L.-T. et al. Rheumatoid arthritis was negatively associated with Alzheimer’s disease: a population-based case-control study. PLoS ONE 11, e0168106 (2016).
Li, G.-S. et al. Rheumatoid arthritis is a protective factor against Alzheimer’s disease: a bidirectional two-sample Mendelian randomization study. Inflammopharmacology 32, 863–871 (2024).
Kiyota, T. et al. Granulocyte-macrophage colony-stimulating factor neuroprotective activities in Alzheimer’s disease mice. J. Neuroimmunol. 319, 80–92 (2018).
Ahmed, M. M. et al. The innate immune system stimulating cytokine GM-CSF improves learning/memory and interneuron and astrocyte brain pathology in Dp16 Down syndrome mice and improves learning/memory in wild-type mice. Neurobiol. Dis. 168, 105694 (2022).
Potter, H. et al. Safety and efficacy of sargramostim (GM-CSF) in the treatment of Alzheimer’s disease. Alzheimers Dement. 7, e12158 (2021).
Balusu, S. et al. MEG3 activates necroptosis in human neuron xenografts modeling Alzheimer’s disease. Science 381, 1176–1182 (2023).
Geula, C. et al. Aging renders the brain vulnerable to amyloid β-protein neurotoxicity. Nat. Med. 4, 827–831 (1998).
Jin, M. et al. Type-I-interferon signaling drives microglial dysfunction and senescence in human iPSC models of Down syndrome and Alzheimer’s disease. Cell Stem Cell 29, 1135–1153 (2022).
Martini, A. C. et al. Distribution of microglial phenotypes as a function of age and Alzheimer’s disease neuropathology in the brains of people with Down syndrome. Alzheimers Dement. 12, e12113 (2020).
Van Hove, H. et al. A single-cell atlas of mouse brain macrophages reveals unique transcriptional identities shaped by ontogeny and tissue environment. Nat. Neurosci. 22, 1021–1035 (2019).
Sun, N. et al. Human microglial state dynamics in Alzheimer’s disease progression. Cell 186, 4386–4403 (2023).
Li, J.-Y. et al. Lewy bodies in grafted neurons in subjects with Parkinson’s disease suggest host-to-graft disease propagation. Nat. Med. 14, 501–503 (2008).
Hoban, D. B. et al. Impact of α-synuclein pathology on transplanted hESC-derived dopaminergic neurons in a humanized α-synuclein rat model of PD. Proc. Natl Acad. Sci. USA 117, 15209–15220 (2020).
Li, W. et al. Extensive graft-derived dopaminergic innervation is maintained 24 years after transplantation in the degenerating parkinsonian brain. Proc. Natl Acad. Sci. USA 113, 6544–6549 (2016).
Castellano, J. M. et al. Human umbilical cord plasma proteins revitalize hippocampal function in aged mice. Nature 544, 488–492 (2017).
Jaiswal, S. et al. Age-related clonal hematopoiesis associated with adverse outcomes. N. Engl. J. Med. 371, 2488–2498 (2014).
Steensma, D. P. et al. Clonal hematopoiesis of indeterminate potential and its distinction from myelodysplastic syndromes. Blood 126, 9–16 (2015).
Vieira, R. et al. Young glial progenitor cells competitively replace aged and diseased human glia in the adult chimeric mouse brain. Nat. Biotechnol. 42, 719–730 (2024).
Mishra, P. et al. Rescue of Alzheimer’s disease phenotype in a mouse model by transplantation of wild-type hematopoietic stem and progenitor cells. Cell Rep. 42, 112956 (2023).
Xu, Z. et al. Efficient strategies for microglia replacement in the central nervous system. Cell Rep. 32, 108041 (2020).
Yoo, Y., Neumayer, G., Shibuya, Y., Marc-Daniel Mader, M. & Wernig, M. A cell therapy approach to restore microglial Trem2 function in a mouse model of Alzheimer’s disease. Cell Stem Cell 30, 1043–1053 (2023).
Chadarevian, J. P. et al. Engineering an inhibitor-resistant human CSF1R variant for microglia replacement. J. Exp. Med. 220, e20220857 (2023).
Liou, J-J. et al. A neuropathology case report of a woman with Down syndrome who remained cognitively stable: implications for resilience to neuropathology. Alzheimers Dement. 21, e14479 (2025).
Papetti, A. V., Jin, M., Ma, Z., Stillitano, A. C. & Jiang, P. Chimeric brain models: unlocking insights into human neural development, aging, diseases, and cell therapies. Neuron 113, 2230–2250 (2025).
Mancuso, R. et al. Stem-cell-derived human microglia transplanted in mouse brain to study human disease. Nat. Neurosci. 22, 2111–2116 (2019).
McQuade, A. et al. Gene expression and functional deficits underlie TREM2-knockout microglia responses in human models of Alzheimer’s disease. Nat. Commun. 11, 5370 (2020).
Jin, M. et al. Co-transplantation-based human-mouse chimeric brain models to study human glial-glial and glial-neuronal interactions. Preprint at bioRxiv https://doi.org/10.1101/2024.07.03.601990 (2024).
Gibson, D. G. et al. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 6, 343–345 (2009).
Li, S., Zhang, A., Xue, H., Li, D. & Liu, Y. One-step piggyBac transposon-based CRISPR/Cas9 activation of multiple genes. Mol. Ther. Nucleic Acids 8, 64–76 (2017).
Cong, L. et al. Multiplex genome engineering using CRISPR/Cas systems. Science 339, 819–823 (2013).
Ran, F. A. et al. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 8, 2281–2308 (2013).
Pille, M. et al. Gene editing-based targeted integration for correction of Wiskott-Aldrich syndrome. Mol. Ther. Methods Clin. Dev. 32, 101208 (2024).
Guschin, D. Y. et al. A rapid and general assay for monitoring endogenous gene modification. Methods Mol. Biol. 649, 247–256 (2010).
Xue, H., Wu, J., Rao, M. S. & Liu, Y. Genetic modification in human pluripotent stem cells by homologous recombination and CRISPR/Cas9 system. Methods Mol. Biol. 1307, 173–190 (2016).
Li, S. et al. Efficient generation of hiPSC neural lineage specific knockin reporters using the CRISPR/Cas9 and Cas9 double nickase system. J. Vis. Exp. 99, e52539 (2015).
Macarthur, C. C. et al. Chromatin insulator elements block transgene silencing in engineered human embryonic stem cell lines at a defined chromosome 13 locus. Stem Cells Dev. 21, 191–205 (2012).
Chen, C. et al. Role of astroglia in Down’s syndrome revealed by patient-derived human-induced pluripotent stem cells. Nat. Commun. 5, 4430 (2014).
Xu, R. et al. OLIG2 drives abnormal neurodevelopmental phenotypes in human iPSC-based organoid and chimeric mouse models of Down syndrome. Cell Stem Cell 24, 908–926 (2019).
Haenseler, W. et al. A highly efficient human pluripotent stem cell microglia model displays a neuronal-co-culture-specific expression profile and inflammatory response. Stem Cell Rep. 8, 1727–1742 (2017).
Jin, M., Alam, M. M., Liu, A. Y.-C. & Jiang, P. Rag2−/− accelerates lipofuscin accumulation in the brain: implications for human stem cell brain transplantation studies. Stem Cell Reports 17, 2381–2391 (2022).
Jin, M., Ma, Z. & Jiang, P. Generation of iPSC-based human-mouse microglial brain chimeras to study senescence of human microglia. STAR Protoc. 3, 101847 (2022).
Xu, R. et al. Human iPSC-derived mature microglia retain their identity and functionally integrate in the chimeric mouse brain. Nat. Commun. 11, 1577 (2020).
Sanchez-Mejias, E. et al. Soluble phospho-tau from Alzheimer’s disease hippocampus drives microglial degeneration. Acta Neuropathol. 132, 897–916 (2016).
Iba, M. et al. Synthetic tau fibrils mediate transmission of neurofibrillary tangles in a transgenic mouse model of Alzheimer’s-like tauopathy. J. Neurosci. 33, 1024–1037 (2013).
Peeraer, E. et al. Intracerebral injection of preformed synthetic tau fibrils initiates widespread tauopathy and neuronal loss in the brains of tau transgenic mice. Neurobiol. Dis. 73, 83–95 (2015).
Boluda, S. et al. Differential induction and spread of tau pathology in young PS19 tau transgenic mice following intracerebral injections of pathological tau from Alzheimer’s disease or corticobasal degeneration brains. Acta Neuropathol. 129, 221–237 (2015).
Jin, M. et al. Interleukin-17 and Th17 Lymphocytes directly impair motoneuron survival of wildtype and FUS-ALS mutant human iPSCs. Int. J. Mol. Sci. 22, 8042 (2021).
Flores-Aguilar, L. et al. Evolution of neuroinflammation across the lifespan of individuals with Down syndrome. Brain 143, 3653–3671 (2020).
Legland, D., Arganda-Carreras, I. & Andrey, P. MorphoLibJ: integrated library and plugins for mathematical morphology with ImageJ. Bioinformatics 32, 3532–3534 (2016).
Miller, S. A. et al. LSD1 and aberrant DNA methylation mediate persistence of enteroendocrine progenitors that support BRAF-mutant colorectal cancer. Cancer Res. 81, 3791–3805 (2021).
Acknowledgements
We thank the UCI-ADRC and the BrightFocus Foundation for providing us with the DSAD and Control human brain tissues. We thank P. Xie (Rutgers University) for assistance with flow cytometry and K. Kwan (Rutgers University) for help with scRNA-seq library preparation. We also thank A. Kumar and K. Aluru from the Jiang laboratory for their assistance with immunohistochemistry, and F. Chanut for editorial assistance. This work was in part supported by grants from the NIH (grant nos. R01NS102382, R01NS122108 and R01AG073779 to P.J.). M.J. was supported by a postdoctoral fellowship award from the New Jersey Department of Health (no. CAUT24DFP004). A.V.P. was supported by a graduate trainee T32 fellowship award from the Training in Translating Neuroscience to Therapies program at Rutgers University (no. T32NS115700). Y.L. was supported by NIH grant no. R01NS110707. Additional support came from the NIH (grant nos. R01AG064579 and RF1NS128800), the JSRM Foundation, the BrightFocus Foundation (BFF17-0008) and the Alzheimer’s Association to S.F, as well as UCI-NIH funding (NIH/National Institute of Aging grant no. P30AG066519) supporting the UCI-ADRC.
Author information
Authors and Affiliations
Contributions
M.J. and P.J. designed the experiments and interpreted the data. M.J. carried out most of the experiments with technical assistance from H.Z. and R.D. Z.M. performed the RNA-seq data analyses and assisted with the interpretation of the sequencing data. Yan Liu and H.Y. performed the donor assignment of the microglia scRNA-seq analyses. R.D. performed the electrophysiological recordings. J.P. and E.H. prepared the human brain tissue extracts. Ying Liu and H.X. generated the CSF2RB A455D DS and CAGG hPSC lines. R.K. assisted with immunostaining and the Imaris analysis. A.V.P. assisted with the flow cytometry experiment. S.F. and E.H. provided critical suggestions to the study. P.J. conceived the study, directed the project and wrote the manuscript together with M.J. and input from all coauthors.
Corresponding author
Ethics declarations
Competing interests
P.J. and M.J. have filed a patent related to this work (US patent application no. 63/563,637). The other authors declare no competing interests.
Peer review
Peer review information
Nature Neuroscience thanks Li Gan and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Generation and characterization of A455D hiPSC.
a-b, Schematic of targeting CSF2RB WT or A455D mutant to intron 10. Donor sequences of both WT and A455D mutant are mammalian codon optimized. P2A is the self-cleaving peptide that allows for simultaneous, separate protein expression of EGFP (Panel a) or mRuby (Panel b) cassette. Puro, puromycin resistance fragment; SA, splice acceptor. c-f, PCR identification and Sanger sequencing verification of targeted hiPSC clones. Genomic DNA extracted from single clones was examined by PCR for identification of correctly targeted alleles (c, d), which were subsequently verified by Sanger sequencing (e, f).
Extended Data Fig. 2 scRNA-seq analysis of DS-WT and DS-A455D microglia in response to DSAD-Tau S1 fraction in chimeric mice.
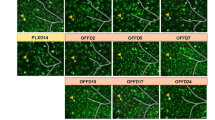
a-b, Representative images of hTMEM119+hN+ and Ki67+hN+ in DS-WT and DS-A455D chimeric mice. Scale bar: 20 μm and 10 μm in the original and enlarged images, respectively. c-d, Quantification of the percentage of hTMEM119+ and Ki67+ in hN+ cells from DS-WT and DS-A455D chimeric mice (Data represent both sexes combined, n = 4 mice per group), Student’s t-test, NS, not significant. Data are presented as mean ± SEM. e, Flow cytometry analysis of p-STAT5 expression in DS-WT and DS-A455D groups. f, Quantification of p-STAT5 expression in DS-WT and DS-A455D groups (Data represent both sexes combined, n = 4 mice per group). Data were analyzed by a two-tailed unpaired t-test, #P = 0.0286. Data are presented as mean ± SEM. g, Representative images of sagittal brain sections showing the distribution of AT8+ puncta at two months post-injection. Scale bar: 200 and 50 μm in the original and enlarged images, respectively. h, Representative images of immunohistochemistry staining with AT8 from control-Tau and DSAD-Tau injected chimeric mouse. Scale bar: 10 and 5 μm in the original and enlarged images, respectively. i, Quantification of the percentage of AT8+ cells in the hippocampus (Data represent both sexes combined, n = 4 mice per group). Arrowheads indicate AT8+ staining. Data were analyzed by a two-tailed unpaired t-test, *P < 0.05. Data are presented as mean ± SEM. j, Representative images of Ki67+hN+ in DS-WT-DSAD-Tau and DS-A455D-DSAD-Tau chimeric mice. Scale bar: 20 μm and 10 μm in the original and enlarged images, respectively. k, Quantification of the percentage of Ki67 in hN+ cells from DS-WT-DSAD-Tau and DS-A455D-DSAD-Tau chimeric mice (Data represent both sexes combined, n = 4 mice per group), Data were analyzed by a two-tailed unpaired t-test, #P < 0.05. Data are presented as mean ± SEM.
Extended Data Fig. 3 scRNA-seq analysis of DS-WT and DS-A455D microglia in response to DSAD-Tau S1 fraction in chimeric mice.
a, A schematic diagram showing the design of the scRNA-seq experiment. b, Violin plots representing the quality control parameters (gene count, UMI counts, and mitochondrial gene percentage) of the scRNA-seq dataset in Cont- and DSAD-Tau-treated groups. Violin plots showing the distribution of detected RNA features (nFeature_RNA) across different clusters (0-9). c, Split UMAP plots showing microglial subclusters (clusters 0-9) from 4 experimental groups. d, Heatmap showing cluster-specific marker gene expression. e, Violin plot showing 20 canonical microglia marker gene expression levels in each microglial subcluster. f, Heatmap showing cosine similarity scores calculated between clusters. g, Violin plot showing canonical DAM marker gene expression levels in each microglial subcluster. h, UMAP plots colored by features of DAM scores, phagocytic microglia scores, FTL, and FTH1 expression. Schematics in a was created using Biorender. Jin, M. (2025) https://BioRender.com/w18g951. i, Barplot showing the proportion of cell cycle phases between clusters. j, Barplots showing the proportion of cells in each cluster between experimental groups. k, UMAP representation of all inferred trajectories of DS-A455D microglia in response to control-Tau and DSAD-Tau. l, Dynamic plots showing additional selected gene expression profiles across inferred pseudotime in lineage 1 and lineage 3. Dots in the plots are cells colored by microglial subclusters.
Extended Data Fig. 4 scRNA-seq analysis of DS-A455D microglia in response to DSAD-Tau S1 fraction.
a, Schematic of the scRNA-seq experimental design for two 10-month-old samples. b, Violin plots showing quality control parameters for the scRNA-seq dataset. c, Bar plots showing the distribution of cells across clusters in each sample. d, UMAP plot of microglial clusters (clusters 0–7) from the two 10-month-old samples. e, Dot plot showing the expression levels of human microglial marker genes across clusters. f, Ridgeline plot of DAM score distributions across clusters. g, Heatmap showing cluster similarity between 4-month-old and 10-month-old samples, with Jaccard scores representing the proportion of overlapping genes. Statistical significance of overlap was determined via Fisher’s exact test. h, Scatter plot of significantly differentially expressed genes in each cluster. i, Bar plot of selected protective GO BP terms from enrichment analysis of the top 100 positive significant marker genes (adjusted p-value < 0.05, ranked by average LogFC). j, Bar plots comparing proportions of homeostatic microglia and DAM between 10-month-old and 4-month-old samples. k, GO enrichment analysis of positive marker genes for each cluster, no IFN-related terms are enriched. l, UMAP plots comparing the density of IFN α/β-responsive gene-expressing cells between 10-month-old and 4-month-old samples. Schematics in a was created using Biorender. Jin, M. (2025) https://BioRender.com/59efcbt.
Extended Data Fig. 5 CSF2RB A455D mutation protects DS microglia against senescence.
a, Ridgeline and box plots showing the senescence score in DS-WT and DS-A455D microglia under control-Tau and DSAD-Tau conditions. Wilcoxon test, ***P < 0.001. b, GSEA plot showing enrichment of interferon genes in DS-A455D-DSAD-Tau and DS-WT-DSAD-Tau groups. c, qPCR analysis of IRF8 and IFITM3 mRNA expression in chimeric mice at month 4 (n = 4-5 mice per group). Student’s t-test, *P = 0.0286, **P = 0.0079. Data are presented as mean ± SEM. d, Representative images of anti-hCD45 (green) and anti-p21 staining (red) in DS-WT-DSAD-Tau and DS-A455D-DSAD-Tau groups; Arrows indicate p21+ and/or hCD45+ staining. Scale bar, 10 and 5 μm in the original and enlarged images, respectively. e, Quantification of the percentage of p21 in hCD45+ cells (Data represent both sexes combined, n = 4 mice per group). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286. Data are presented as mean ± SEM. f, Representative raw fluorescence super-resolution and 3D surface rendered images showing images of colocalization of LAMP1+ and LC3B in hCD45+ microglia in 4-month-old DS-WT-DSAD-Tau and DS-A455D-DSAD-Tau chimeric mice. Scale bars: 5 μm and 3 μm in the original and enlarged images, respectively. g, Quantification of LAMP1+ puncta in hCD45+ microglia in 4-month-old DS-WT-DSAD-Tau and DS-A455D-DSAD-Tau chimeric mice (Data represent both sexes combined, n = 5 mice per group). Data were analyzed by a two-tailed unpaired t-test, **P = 0.0079. Data are presented as mean ± SEM. h, Quantification of LC3B+ puncta in hCD45+ microglia in 4-month-old DS-WT-DSAD-Tau and DS-A455D-DSAD-Tau chimeric mice (Data represent both sexes combined, n = 5 mice per group). Data were analyzed by a two-tailed unpaired t-test, **P = 0.0079. Data are presented as mean ± SEM. i, Quantification of colocalization of LAMP1+ and LC3B in hCD45+ microglia in 4-month-old DS-WT-DSAD-Tau and DS-A455D-DSAD-Tau chimeric mice (Data represent both sexes combined, n = 5 mice per group). Data were analyzed by a two-tailed unpaired t-test, **P = 0.0079. Data are presented as mean ± SEM.
Extended Data Fig. 6 AC-4–130 treatment abolishes the protective effects of the CSF2RB A455D mutation on human microglia.
a, Representative images showing colocalization of hCD45+ and Ferritin+ staining in DS-A455D-DSAD-Tau+vehicle and DS-A455D-DSAD-Tau+AC-4-130 groups. Arrows indicate hCD45+ and/or Ferritin+ staining. Scale bar: 20 μm. Representative images of IBA+/hN+ human microglia in DS-A455D-DSAD-Tau+vehicle and DS-A455D-DSAD-Tau+AC-4-130 groups. Scale bars: 5 μm b, Quantification of microglia volumes (Data represent both sexes combined, n = 5 mice per group). Data were analyzed by a two-tailed unpaired t-test, **P = 0.0079. Data are presented as mean ± SEM. c, Quantification of the process length, soma size, and soma size/process length (n = 5 mice per group). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0317, **P = 0.0079, NS, not significant. Data are presented as mean ± SEM. d, Quantification of the percentage of Ferritin in hCD45+ microglia cells (Data represent both sexes combined, n = 5 mice per group). Data were analyzed by a two-tailed unpaired t-test, * P = 0.0079. Data are presented as mean ± SEM. e, Representative raw fluorescence super-resolution and 3D surface rendered images showing images of colocalization of LAMP1+ and LC3B+ in hCD45+ microglia in 4-month-old DS-A455D chimeric mice. Scale bars: 5 μm and 2 μm in the original and enlarged images, respectively. f, Quantification of LAMP1+ puncta in hCD45+ microglia in 4-month-old DS-A455D chimeric mice (Data represent both sexes combined, n = 5 mice per group). Data were analyzed by a two-tailed unpaired t-test, ## P = 0.0022. Data are presented as mean ± SEM. g, Quantification of LC3B+puncta in hCD45+ microglia in 4-month-old DS-A455D chimeric mice (n = 5 mice per group). Data were analyzed by a two-tailed unpaired t-test, ## P = 0.0022. Data are presented as mean ± SEM. h, Quantification of LAMP1+ and LC3B+ colocalization in hCD45+ microglia in 4-month-old DS-A455D chimeric mice (Data represent both sexes combined, n = 5 mice per group). Data were analyzed by a two-tailed unpaired t-test, ## P = 0.0022. Data are presented as mean ± SEM. i, Representative image of LTP recording slice from transverse sagittal brain sections showing the distribution of transplanted microglia in 4-month-old chimeric mouse brains. Scale bar: 300 μm.
Extended Data Fig. 7 CSF2RB A455D mutation resilience is independent of trisomy 21.
a, Quantification of CD235+, CD43+, and Ki67+ PMPs derived from the control-WT and control-A455D hiPSC lines (n = 4, each experiment was repeated four times). Student’s t-test, NS, not significant. Data are presented as mean ± SEM. b, The A455D rate in control-WT and control-A455D PMP (n = 3, each experiment was repeated three times). Data were analyzed by a two-tailed unpaired t-test, ****P < 0.0001. Data are presented as mean ± SEM. c, Linear regression and Spearman’s correlation test of RNA-seq data between control-WT and control-A455D PMP at the full transcriptome level. d, Volcano plots comparing control-WT and control-A455D PMP. e, Flow cytometry analysis of p-STAT5 level in control-WT and control-A455D PMP. f, Quantification of p-STAT5 level in control-WT and control-A455D PMP (n = 5, each experiment was repeated five times; Geo mean: Geometric mean). Data were analyzed by a two-tailed unpaired t-test, ##P = 0.0079. Data are presented as mean ± SEM. g, Quantification of IBA-1+ and CD45+ microglia derived from the control-WT and control-A455D hiPSC lines (n = 4, each experiment was repeated four times). Data were analyzed by a two-tailed unpaired t-test, NS, not significant. Data are presented as mean ± SEM. h, Flow cytometry analysis of p-STAT5 level in control-WT and control-A455D microglia. i, Quantification of p-STAT5 expression in control-WT and control-A455D microglia (n = 5, each experiment was repeated five times; Geo mean: Geometric mean). Data were analyzed by a two-tailed unpaired t-test, ##P = 0.0079. Data are presented as mean ± SEM. j, Quantitative analysis of cell viability after Tau treatment (n = 6, each experiment was repeated six times). Data were analyzed by a one-way ANOVA followed by a Tukey multiple comparison test, *P = 0.0358 and 0.0020, ****P < 0.0001. Data are presented as mean ± SEM. k, Representative live-cell imaging of microglia from control-WT and control-A455D iPSCs in the phagocytosis of zymosan particles. Bright-field images are overlaid for reference. Scale bars: 100 μm. l, Quantification of the proportion of microglia with zymosan particles (n = 5, each experiment was repeated five times). Data were analyzed by a two-tailed unpaired t-test, ##P = 0.0079. Data are presented as mean ± SEM. m, Representative images of hTMEM119+hN+ and Ki67+hN+ in control-WT and control-A455D chimeric mice. Scale bar: 20 μm. n, Quantification of the percentage of hTMEM119 in hN+ cells from control-WT and control-A455D chimeric mice (n = 4 mice per group), Student’s t-test, NS, not significant. Data are presented as mean ± SEM. o, Quantification of the percentage of Ki67 in hN+ cells from control-WT and control-A455D chimeric mice (Data represent both sexes combined, n = 4 mice per group), Data were analyzed by a two-tailed unpaired t-test, NS, not significant. Data are presented as mean ± SEM. p, Flow cytometry analysis of p-STAT5 level in control-WT and control-A455D microglia. q, Quantification of p-STAT5 expression in control-WT and control-A455D microglia (Data represent both sexes combined, n = 4 mice per group; Geo mean: Geometric mean). Data were analyzed by a two-tailed unpaired t-test, #P = 0.0286. Data are presented as mean ± SEM. r, Representative images of Ki67+ hN+ in control-WT-DSAD-Tau and control-A455D DSAD-Tau chimeric mice. Scale bars: 20 μm and 10 μm in the original and enlarged images, respectively. s, Quantification of the percentage of Ki67+ in hN+ cells from control-WT-DSAD-Tau and control-A455D DSAD-Tau chimeric mice (Data represent both sexes combined, n = 5 mice per group), Data were analyzed by a two-tailed unpaired t-test, ##P = 0.0079. Data are presented as mean ± SEM.
Extended Data Fig. 8 The influence of CSF2RB A455D mutation in Control microglia.
a, Representative images showing colocalization of hCD45+ and Ferritin+ staining in CAGG-WT-DSAD-Tau and CAGG-A455D-DSAD-Tau groups. Arrows indicate hCD45+ and/or Ferritin+ staining. Scale bar: 20 μm. Representative images of IBA+/hN+ human microglia in control-Tau and DSAD-Tau groups. Scale bars: 5 μm b, Quantification of microglia volumes in CAGG-WT and CAGG-A455D microglia following control and DSAD Tau injection (Data represent both sexes combined, n = 4 mice per group). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286. Data are presented as mean ± SEM. c, Quantification of the process length, soma size, and soma size/process length (Data represent both sexes combined, n = 4 mice per group). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286, NS, not significant. Data are presented as mean ± SEM. d, Quantification of the percentage of Ferritin in hCD45+ cells (Data represent both sexes combined, n = 4 mice per group). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286. Data are presented as mean ± SEM. e, Representative image of LTP recording slice from transverse sagittal brain sections showing the distribution of transplanted microglia in 6-month-old chimeric mice. Scale bar: 300 μm. f, Whole transcriptomic level linear regression and Spearman’s correlation analysis of gene expression levels from bulk RNA-seq data, comparing GFP+ and GFP− WT/A455D PMPs.
Extended Data Fig. 9 scRNA-seq analysis of control A455D microglia and control WT microglia exposed to DSAD-Tau S1 fraction.
a, Representative images from sagittal brain sections showing the distribution of transplanted CAGG-A455D and control-WT at the age of 6-month-old chimeric mice. Scale bar: 300 μm. b, Quantification of the percentage of control-WT and CAGG-A455D microglia in total microglia cells from 6-month-old chimeric mice (Data represent both sexes combined, n = 4 mice per group), Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286, NS, not significant. Data are presented as mean ± SEM. c, Representative raw fluorescence super-resolution and 3D surface rendered images showing images of colocalization of LC3B+ in hCD45+ microglia and GFP+ microglia in 6-month-old CAGG-A455D-control-WT chimeric mice receiving injection of DSAD Tau at the age of 8 weeks. Scale bars: 5 μm and 2 μm in the original and enlarged images, respectively. d, Quantification of LC3B+ puncta in control-WT and CAGG-A455D microglia following DSAD-Tau injection (Data represent both sexes combined, n = 4 mice per group). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286. Data are presented as mean ± SEM. e, Representative raw fluorescence super-resolution and 3D surface rendered images showing images of colocalization of CD68+ in hTMEM119+ microglia and GFP+ microglia in 6-month-old CAGG-A455D-control-WT chimeric mice receiving injection of DSAD Tau at the age of 8 weeks. Scale bars: 5 μm and 2 μm in the original and enlarged images, respectively. f, Quantification of CD68+ puncta in control-WT and CAGG-A455D microglia following DSAD-Tau injection (Data represent both sexes combined, n = 4 mice per group). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286. Data are presented as mean ± SEM. g, Representative images of Dcx and NeuN staining in 6-month-old chimeric mice. Scale bars: 20 μm. h, Quantification of Dcx+ cells (Data represent both sexes combined, n = 4 mice per group). Data were analyzed by a two-tailed unpaired t-test, NS, not significant. Data are presented as mean ± SEM. i, Scatter plot showing significantly (adj. p-value < 0.05) differentially expressed genes between two comparison groups ranked by average logFC. Genes located in the third quadrant are those both downregulated in DS-A455D-DSAD-tau compared to DS-control-DSAD-tau and downregulated in WT-A455D-DSAD-tau relative to WT-control-DSAD-tau. j-k, Reactome and GO-BP enrichment analyses of the co-downregulated genes. l, Violin plots showing the senescence score of co-transplantation each group. Wilcoxon test, n.s.: not significant, ***P < 0.001.
Extended Data Fig. 10 Control A455D microglia replace Control WT microglia in the tau pathology model.
a-b, Representative image of brain sections from animals (6 months or 10 months of age) expressing P301L-Tau. Scale bar, 500 μm and 20 μm respectively. c-e, Representative images of panel antibodies targeting tau forms, including total transgenic tau (HT7), phosphor-tau (AT8 and PHF1) in the hippocampus of chimeric mice brains (6 months of age) expressing P301L-Tau. Scale bar, 20 μm. f, Representative images showing colocalization of hTMEM119+ microglia and P301L-Tau in 6-month-old chimeric mice. Scale bar: 50 μm. g, Representative images showing colocalization of hCD45+ GFP+ and hCD45+ GFP− staining in 6-month-old chimeric mice. Scale bar: 20 μm. h, Quantification of the percentage of CAGG-WT (hCD45+ GFP+) and Control-A455D (hCD45+ GFP−) microglia in total hCD45+ microglia cells from 6-month-old chimeric mice (Data represent both sexes combined, n = 4 mice per group), Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286, NS, not significant. Data are presented as mean ± SEM. i, Representative raw fluorescence super-resolution and 3D surface rendered images showing images of hCD45+GFP+ microglia and hCD45+GFP− microglia in 6-month-old chimeric mice receiving injection of AAV-P301L-Tau. Scale bars: 5 μm and 2 μm in the original and enlarged images, respectively. j-l, Quantification of the process length, soma size, and soma size/process length (Data represent both sexes combined, n = 4 mice per group). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286. Data are presented as mean ± SEM. m, Quantification of the microglia volumes of CAGG-WT and Control-A455D in the AAV-P301L-Tau group (Data represent both sexes combined, n = 4 mice). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286. Data are presented as mean ± SEM. n, Representative raw fluorescence super-resolution and 3D surface rendered images showing colocalization of hCD45+ GFP− microglia and GFP+ staining in 6-month-old chimeric mice receiving injection of AAV-P301L-Tau at the age of 8 weeks. Scale bar: 5 μm, 3 μm, and 2μm in the original and enlarged images, respectively. o, Quantification of the GFP volumes of CAGG-WT and control-A455D in the AAV-P301L-Tau group (Data represent both sexes combined, n = 4 mice). Data were analyzed by a two-tailed unpaired t-test, *P = 0.0286. Data are presented as mean ± SEM.
Supplementary information
Supplementary Information (download PDF )
Supplementary Fig. 1 and Tables 1–12.
Supplementary Tables 6–11 (download XLSX )
Supplementary Tables 6–11.
Source data
Source Data (download XLSX )
Statistical Source Data for Figs. 1–7 and Extended Data Figs. 1–10.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Jin, M., Ma, Z., Dang, R. et al. A myeloid trisomy 21-associated gene variant is protective from Alzheimer’s disease. Nat Neurosci 29, 25–39 (2026). https://doi.org/10.1038/s41593-025-02117-8
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41593-025-02117-8
