
Abstract
N-Lactoyl-phenylalanine (Lac-Phe) is a lactate-derived circulating metabolite that reduces feeding and obesity, but the molecular mechanisms that underlie the metabolic benefits of Lac-Phe remain unknown. Here we show that Lac-Phe directly inhibits hypothalamic neurons that express Agouti-related protein (AgRP), resulting in an indirect activation of anorexigenic neurons in the paraventricular nucleus of the hypothalamus (PVH). Both AgRP inhibition and PVH activation are required to mediate Lac-Phe-induced hypophagia. Lac-Phe-mediated inhibition of AgRP neurons occurs through activation of the ATP-sensitive potassium (KATP) channel, whereas inhibition of the KATP channel blunts the effects of Lac-Phe to suppress feeding. Together, these results reveal the molecular and neurobiological mechanisms by which Lac-Phe mediates metabolic improvements and suggest this exercise-induced metabolite might have therapeutic benefits in various human diseases.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Data availability
Numerical source data are published alongside the article. Source data are provided with this paper.
References
Eriksson, K. F. & Lindgarde, F. Prevention of type 2 (non-insulin-dependent) diabetes mellitus by diet and physical exercise. The 6-year Malmö feasibility study. Diabetologia 34, 891–898 (1991).
Pan, X. R. et al. Effects of diet and exercise in preventing NIDDM in people with impaired glucose tolerance. The Da Qing IGT and Diabetes Study. Diabetes Care 20, 537–544 (1997).
Rejeski, W. J. et al. Lifestyle change and mobility in obese adults with type 2 diabetes. N. Engl. J. Med. 366, 1209–1217 (2012).
Rawshani, A. et al. Risk factors, mortality, and cardiovascular outcomes in patients with type 2 diabetes. N. Engl. J. Med. 379, 633–644 (2018).
Helmrich, S. P., Ragland, D. R., Leung, R. W. & Paffenbarger, R. S. Jr. Physical activity and reduced occurrence of non-insulin-dependent diabetes mellitus. N. Engl. J. Med. 325, 147–152 (1991).
Li, V. L. et al. An exercise-inducible metabolite that suppresses feeding and obesity. Nature 606, 785–790 (2022).
Hoene, M. et al. Exercise-induced N-lactoylphenylalanine predicts adipose tissue loss during endurance training in overweight and obese humans. Metabolites 13, 15 (2022).
Xiao, S. et al. Lac-Phe mediates the effects of metformin on food intake and body weight. Nat. Metab. 6, 659–669 (2024).
Scott, B. et al. Metformin and feeding increase levels of the appetite-suppressing metabolite Lac-Phe in humans. Nat. Metab. 6, 651–568 (2024).
Rogers, R. S. et al. Circulating N-lactoyl-amino acids and N-formyl-methionine reflect mitochondrial dysfunction and predict mortality in septic shock. Metabolomics 20, 36 (2024).
Jansen, R. S. et al. N-lactoyl-amino acids are ubiquitous metabolites that originate from CNDP2-mediated reverse proteolysis of lactate and amino acids. Proc. Natl Acad. Sci. USA 112, 6601–6606 (2015).
Sutton, A. K., Myers, M. G. Jr. & Olson, D. P. The role of PVH circuits in leptin action and energy balance. Annu. Rev. Physiol. 78, 207–221 (2016).
Cheng, W. et al. Hindbrain circuits in the control of eating behaviour and energy balance. Nat. Metab. 4, 826–835 (2022).
Guenthner, C. J., Miyamichi, K., Yang, H. H., Heller, H. C. & Luo, L. Permanent genetic access to transiently active neurons via TRAP: targeted recombination in active populations. Neuron 78, 773–784 (2013).
DeNardo, L. A. et al. Temporal evolution of cortical ensembles promoting remote memory retrieval. Nat. Neurosci. 22, 460–469 (2019).
Beier, K. T. et al. Circuit architecture of VTA dopamine neurons revealed by systematic input–output mapping. Cell 162, 622–634 (2015).
Morton, G. J., Cummings, D. E., Baskin, D. G., Barsh, G. S. & Schwartz, M. W. Central nervous system control of food intake and body weight. Nature 443, 289–295 (2006).
Hahn, T. M., Breininger, J. F., Baskin, D. G. & Schwartz, M. W. Coexpression of Agrp and NPY in fasting-activated hypothalamic neurons. Nat. Neurosci. 1, 271–272 (1998).
Yang, D. et al. Phosphorylation of pyruvate dehydrogenase inversely associates with neuronal activity. Neuron 112, 959–971.e8 (2024).
Xu, J. et al. Genetic identification of leptin neural circuits in energy and glucose homeostases. Nature 556, 505–509 (2018).
He, Y. et al. Human loss-of-function variants in the serotonin 2C receptor associated with obesity and maladaptive behavior. Nat. Med. 28, 2537–2546 (2022).
D’Agostino, G. et al. Nucleus of the solitary tract serotonin 5-HT2C receptors modulate food intake. Cell Metab. 28, 619–630.e5 (2018).
He, Z. et al. Cellular and synaptic reorganization of arcuate NPY/AgRP and POMC neurons after exercise. Mol. Metab. 18, 107–119 (2018).
Miletta, M. C. et al. AgRP neurons control compulsive exercise and survival in an activity-based anorexia model. Nat. Metab. 2, 1204–1211 (2020).
Landry, T. et al. Exercise increases NPY/AgRP and TH neuron activity in the hypothalamus of female mice. J. Endocrinol. 252, 167–177 (2022).
Landry, T. et al. Energy status differentially modifies feeding behavior and POMCARC neuron activity after acute treadmill exercise in untrained mice. Front. Endocrinol. (Lausanne) 12, 705267 (2021).
Krashes, M. J. et al. Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J. Clin. Invest. 121, 1424–1428 (2011).
Aponte, Y., Atasoy, D. & Sternson, S. M. AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nat. Neurosci. 14, 351–355 (2011).
Krashes, M. J., Shah, B. P., Koda, S. & Lowell, B. B. Rapid versus delayed stimulation of feeding by the endogenously released AgRP neuron mediators GABA, NPY, and AgRP. Cell Metab. 18, 588–595 (2013).
Mishra, I. et al. Protein tyrosine phosphatase receptor delta serves as the orexigenic asprosin receptor. Cell Metab. 34, 549–563.e8 (2022).
Duerrschmid, C. et al. Asprosin is a centrally acting orexigenic hormone. Nat. Med. 23, 1444–1453 (2017).
Feng, B. et al. Asprosin promotes feeding through SK channel-dependent activation of AgRP neurons. Sci. Adv. 9, eabq6718 (2023).
Wang, Q. et al. Arcuate AgRP neurons mediate orexigenic and glucoregulatory actions of ghrelin. Mol. Metab. 3, 64–72 (2014).
Doslikova, B. et al. 5-HT2C receptor agonist anorectic efficacy potentiated by 5-HT1B receptor agonist coapplication: an effect mediated via increased proportion of pro-opiomelanocortin neurons activated. J. Neurosci. 33, 9800–9804 (2013).
Heisler, L. K. et al. Serotonin reciprocally regulates melanocortin neurons to modulate food intake. Neuron 51, 239–249 (2006).
He, Y. et al. 5-HT recruits distinct neurocircuits to inhibit hunger-driven and non-hunger-driven feeding. Mol. Psychiatry 26, 7211–7224 (2021).
Sun, J. et al. Adiponectin potentiates the acute effects of leptin in arcuate Pomc neurons. Mol. Metab. 5, 882–891 (2016).
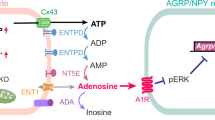
Song, Z. & Routh, V. H. Differential effects of glucose and lactate on glucosensing neurons in the ventromedial hypothalamic nucleus. Diabetes 54, 15–22 (2005).
Parsons, M. P. & Hirasawa, M. ATP-sensitive potassium channel-mediated lactate effect on orexin neurons: implications for brain energetics during arousal. J. Neurosci. 30, 8061–8070 (2010).
Karagiannis, A. et al. Lactateis an energy substrate for rodent cortical neurons and enhances their firing activity. eLife 10, e71424 (2021).
Betley, J. N., Cao, Z. F., Ritola, K. D. & Sternson, S. M. Parallel, redundant circuit organization for homeostatic control of feeding behavior. Cell 155, 1337–1350 (2013).
Wu, Q., Clark, M. S. & Palmiter, R. D. Deciphering a neuronal circuit that mediates appetite. Nature 483, 594–597 (2012).
Madisen, L. et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 13, 133–140 (2010).
Tong, Q., Ye, C. P., Jones, J. E., Elmquist, J. K. & Lowell, B. B. Synaptic release of GABA by AgRP neurons is required for normal regulation of energy balance. Nat. Neurosci. 11, 998–1000 (2008).
Berglund, E. D. et al. Serotonin 2C receptors in pro-opiomelanocortin neurons regulate energy and glucose homeostasis. J. Clin. Invest. 123, 5061–5070 (2013).
Tang, Y. et al. Viral vectors for opto-electrode recording and photometry-based imaging of oxytocin neurons in anesthetized and socially interacting rats. STAR Protoc. 3, 101032 (2022).
Clasadonte, J., Scemes, E., Wang, Z., Boison, D. & Haydon, P. G. Connexin 43-mediated astroglial metabolic networks contribute to the regulation of the sleep–wake cycle. Neuron 95, 1365–1380.e5 (2017).
Plum, L. et al. Enhanced PIP3 signaling in POMC neurons causes KATP channel activation and leads to diet-sensitive obesity. J. Clin. Invest. 116, 1886–1901 (2006).
Cai, X. et al. A D2 to D1 shift in dopaminergic inputs to midbrain 5-HT neurons causes anorexia in mice. Nat. Neurosci. 25, 646–658 (2022).
Acknowledgements
The investigators were supported by grants from the USDA/CRIS (51000-064-01S to Y.X., 3092-51000-062-04(B)S to C.W.), American Heart Association (23POST1030352 to Hailan Liu), National Institutes of Health (F32DK134121 to K.M.C.; R01DK136479 to Y.X., R01DK136526 to J.Z.L., T32GM13854 to V.L.L.), a Bio-X SIGF Graduate Student Fellowship to V.L.L. and Texas Children’s Research Scholar funds to Y.H.
Author information
Authors and Affiliations
Contributions
Hailan Liu, V.L.L., Q.L., Y.H., J.Z.L. and Y.X. conceived of the project and experimental design and wrote the paper. Hailan Liu, V.L.L., Q.L. and Y.H. performed the procedures, data acquisition and analyses. Y. Liu, S.C. and Hueyxian Wong assisted with feeding studies. N.Y. and Yongjie Yang assisted in molecular and cellular experiments. Hesong Liu, X.F., K.M.M., Hueyzhong Wong, M.Y., L.T., J.C.B., Y. Li, M.W., Y.D., Y.S., O.Z.G., Yuxue Yang, J.H., M.E.B. and S.V.J. contributed to the generation of study mice and data discussion. C.W., B.R.A. and D.K. were involved in study design and data discussion.
Corresponding authors
Ethics declarations
Competing interests
V.L.L., Y.H., J.Z.L. and Y.X. are listed as inventors on a patent covering lactoyl amino acids for the treatment of metabolic diseases (E-160-2023-0-PC-01, N-lactoyl-phenylalanine [Lac-Phe] compound derivatives, Stanford University and Baylor College of Medicine). The other authors declare no conflict of interest.
Peer review
Peer review information
Nature Metabolism thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: Christoph Schmitt, in collaboration with the Nature Metabolism team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Lac-Phe does not induce adverse effects.
(a) Three-hour food intake in chow-fed female WT mice (2 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). p = 0.016. (b) Three-hour food intake in HFD-fed female WT mice (3 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). p = 0.0047. (c) Three-hour kaolin intake in male WT mice (3 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). (d) Flavor preference ratio in male WT mice (3 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). (e) Travel distance, velocity, number of entries into the center and time spent in the center during OFT in male WT mice (3 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). (f) Travel distance, velocity, number of entries into the open arms and time spent in the open arms during OFT in male WT mice (3 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). (g) Number of entries into the closed arms and time spent in the closed arms during OFT in male WT mice (3 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). (h) Time spent in immobility in FST in male WT mice (3 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). (i) Sucrose preference ratio in SPT in male WT mice (3 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). N = 8 mice in each group for a-i.Data are mean ± SEM with individual data points in (a-i). Two-sided unpaired t-test was used in (a-i). *, p < 0.05; **, p < 0.01.
Extended Data Fig. 2 Lac-Phe activates NTS and PVH neurons.
(a-b) Representative fluorescent images showing c-Fos (a) and quantification (b) in the NTS in HFD-fed female WT mice (4 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). N = 3 mice in each group. p = 0.00014. (c-d) Representative fluorescent images showing c-Fos (c) and quantification (d) in the PVH in HFD-fed female WT mice. N = 3 mice in each group. p < 0.0001. (e-f) Representative microscopic images showing mCherry signals in the NTS of control and NTSTRAP-Di mice (e) or in the PVH of control and PVHTRAP-Di mice (f). N = 3 biological replicates in each group. (g) A representative current-clamp trace showing effects of CNO (10 μM, 5 s puff) on a mCherry+ PVH neuron in PVHTRAP-Di mice. (h-i) Resting membrane potential (h) and firing frequency (i) of mCherry+ PVH neurons during the baseline and 10 μM CNO treatment. N = 5 neurons from 3 different mice. p = 0.0017 for h; p = 0.0007 for i. Data are mean ± SEM with individual data points in (b, d) and individual data points in (h-i). Two-sided unpaired t-test was used in (b, d); Two-sided paired t-tests were used in (h-i). **, p < 0.01; ***, p < 0.001; ****, p < 0.0001.
Extended Data Fig. 3 Rabies tracing from Lac-Phe-activated PVH neurons.
(a) Representative fluorescent images showing dsRed (red) and DAPI (blue) expression in the PVH, LS, NAcSh, POA, PVT, SON, cHypo, and MVe of male TRAP2 mice with Lac-Phe-activated PVH neurons infected by EnvA-ΔG-Rabies-dsRed. (b) RNAscope images showing tdTomato, Mc4r and Npy1r expression in the PVH of male TRAP2/Rosa26-LSL-tdTomato mice following Lac-Phe TRAP. (c-e) Quantification of the percentage of tdTomato+ cells expressing Mc4r (c), or Npy1r (d), or Mc4r and Npy1r (e). N = 3 mice.
Extended Data Fig. 4 i.p. injection of Lac-Phe increases its concentration in the ARH.
(a-c) Body weight (a), body composition (b), and blood glucose (c) changes in chow- and HFD-fed male mice. N = 19 mice in each group. p < 0.0001 for b; p = 0.0086 for c. (d-e) Lac-Phe concentration in the serum (d) and ARH (e) of chow-fed male mice at 0, 5, 10, 30, 60 and 120 min after an i.p. injection of 50 mg/kg Lac-Phe. N = 4 mice for 0 and 5 min, N = 3 for 10, 30, 60, and 120 min. p = 0.0022 for 0 vs 5; p = 0.0005 for 0 vs 10; p = 0.015 for 0 vs 30 min in d; p = 0.0003 for 0 vs 5; p = 0.0011 for 0 vs 10 min in e. (f-g) Lac-Phe concentration in the serum (f) and ARH (g) of HFD-fed male mice at 0, 5, 10, 30, 60 and 120 min after an i.p. injection of 50 mg/kg Lac-Phe. N = 4 mice for 0, 5 and 10 min, N = 3 for 30, 60, and 120 min. p = 0.0085 for 0 vs 5; p = 0.0035 for 0 vs 10; p = 0.0059 for 0 vs 30, p = 0.0425 for 0 vs 60 min in f; p = 0.0009 for 0 vs 5; p = 0.0184 for 0 vs 10; p = 0.0293 for 0 vs 30, p = 0.0372 for 0 vs 60 min in g. (h-i) Serum (h) and ARH (i) Lac-Phe concentrations in chow and HFD-fed male mice at 0, 5, 10, 30, 60 and 120 min after an i.p. injection of 50 mg/kg Lac-Phe. Note that panels h-i were plotted using the same data from d-g. Data are mean ± SEM with individual data points in (b, d-g) and mean ± SEM in (a, c, h-i). One-way ANOVA analysis was used in (d-g); two-way ANOVA analysis was used in (a-c, h-i). *, p < 0.05; **, p < 0.01; ***, p < 0.001; ****, p < 0.0001.
Extended Data Fig. 5 Lac-Phe activates AgRP neurons during fasting conditions.
(a-b) Temporal changes in the resting membrane potential (a) and firing frequency (b) of AgRP neurons in the ARH in response to 2.5 μM (puff 5 s) lactate or phenylalanine. N = 7 neurons from 3 different mice. (c) Representative current-clamp traces showing effects of vehicle or Lac-Phe (5 μM) on AgRP neurons in overnight fasted male mice. (d-e) Temporal changes in the resting membrane potential (d) and firing frequency (e) of AgRP neurons in male mice following vehicle or Lac-Phe application. N = 6 neurons from 3 different mice for each treatment. p < 0.0001 for d and e. (f) Representative current-clamp traces showing effects of vehicle or Lac-Phe (5 μM) on AgRP neurons in overnight fasted female mice. (g-h) Temporal changes in the resting membrane potential (g) and firing frequency (h) of AgRP neurons in female mice following vehicle or Lac-Phe application. N = 6 neurons from 3 different mice for each treatment. p = 00001 for g, p < 0.0001 for h. Data are mean ± SEM in (a-b, d-e, g-h). Two-way ANOVA analysis followed by post hoc Sidak tests was used in (a-b, d-e, g-h). ***, p < 0.001; ****, p < 0.0001.
Extended Data Fig. 6 Lac-Phe inhibits AgRP neurons.
(a) The experimental design for recording AgRP neuronal activity 1 h after vehicle or Lac-Phe injection. (b) Representative current-clamp traces of AgRP neurons recorded 1 h after vehicle or Lac-Phe injection in male AgRP-IRES-Cre/Rosa26-LSL-tdTomato mice. (c-d) Resting membrane potential (c) and firing frequency (d) of AgRP neurons in male mice following vehicle or Lac-Phe administration. N = 17 (vehicle) or 18 (Lac-Phe) neurons from 3 different mice in each group. p < 0.0001 for c, p = 0.0077 for d. (e) Representative current-clamp traces of AgRP neurons recorded 1 h after vehicle or Lac-Phe injection in female AgRP-IRES-Cre/Rosa26-LSL-tdTomato mice. (f-g) Resting membrane potential (f) and firing frequency (g) of AgRP neurons in female mice following vehicle or Lac-Phe administration. N = 14 (vehicle) or 16 (Lac-Phe) neurons from 3 different mice in each group. p = 0.0011 for f, p = 0.0298 for g. (h) Representative current-clamp traces of AgRP neurons recorded 3 h after vehicle or Lac-Phe injection in female AgRP-IRES-Cre/Rosa26-LSL-tdTomato mice. (i-j) Resting membrane potential (i) and firing frequency (j) of AgRP neurons in female mice following vehicle or Lac-Phe administration. N = 14 neurons from 3 different mice in each group. p = 0.010 for I, p = 0.012 for j. Data are mean ± SEM with individual data points in (c-d, f-g, i-j). Two-sided unpaired t-tests were used in (c-d, f-g, i-j). *, p < 0.05; **, p < 0.01; ****, p < 0.0001.
Extended Data Fig. 7 AgRP neurons mediate exercise-induced hypophagia.
(a) A representative microscopic image showing expression of GCaMP6m in AgRP neurons in the ARH with the fiber photometry probe track. N = 6 biological replicates. (b) Representative microscopic images showing pPDH immunofluorescence (green) in tdTomato-labelled AgRP neurons (red) in male AgRP-IRES-Cre/Rosa26-LSL-tdTomato mice (4 months of age) receiving i.p. injections of vehicle or Lac-Phe (50 mg/kg). (c) Quantifications of pPDH+/tdTomato+ cells. N = 3 mice in each group. p < 0.0001. (d) Quantification for the number of Agrp+ cells in the ARH of control and AgRPDTA mice. N = 9 mice for control, N = 11 mice for AgRPDTA. p < 0.0001. (e) Body weight gain in control and AgRPDTA mice after HFD feeding. N = 9 mice for control, N = 11 mice for AgRPDTA. P = 0.0104. (f) A representative current-clamp trace showing effects of CNO (10 μM, 10 s puff) on an AgRP neurons in AgRPDq mice. (g-h) Resting membrane potential (g) and firing frequency (h) of AgRP neurons during the baseline and 10 μM CNO treatment. N = 7 neurons from 3 different mice. p = 0.0002 for g, p = 0.0017 for h. (i) Representative microscopic images showing c-Fos immunofluorescence (green) in tdTomato-labelled AgRP neurons (red) in male AgRP-IRES-Cre/Rosa26-LSL-tdTomato mice (4 months of age) that were infected with AAV8-FLEX-hM3Dq-mCherry, followed by i.p. injections of saline or CNO (1 mg/kg). (j) Quantifications of c-Fos+/tdTomato+ cells. N = 3 mice in each group. p < 0.0001. Data are mean ± SEM with individual data points in (c-d, j) and mean ± SEM in (e) or individual data points in (g-h). Two-sided unpaired t-tests were used in (c-d, j); two-sided paired t-tests were used in (g-h); two-way ANOVA analysis was used in (e). *, p < 0.05; **, p < 0.01; ***, p < 0.001; ****, p < 0.0001.
Extended Data Fig. 8 Lac-Phe does not activate POMC neurons.
(a) A representative current-clamp trace showing effects of Lac-Phe (5 μM, 5 s puff) on a POMC neuron in POMC-CreERT2/Rosa26-LSL-tdTomato mice. (b-c) Temporal changes in the resting membrane potential (b) and firing frequency (c) of POMC neurons in the ARH in response to vehicle or 5 μM Lac-Phe (puff 5 s). (d) Representative microscopic images showing β-endorphin immunofluorescence (green) in tdTomato-labelled ARH neurons (red) in male TRAP2/Rosa26-LSL-tdTomato mice (3 months of age) following Lac-Phe TRAP. N = 3 mice. Data are mean ± SEM in (b, c). Two-way ANOVA analysis followed by post hoc Sidak tests was used in (b, c).
Extended Data Fig. 9 Lac-Phe enhances KATP channels in AgRP neurons of female mice.
(a) Representative voltage-clamp traces for KATP currents in AgRP neurons of female mice at the baseline (black) or in response to 5 μM Lac-Phe (red) in the absence or presence of tolbutamide. (b) Quantifications of KATP currents in AgRP neurons in response to various treatments as described in (a). N = 6 or 8 neurons from 3 different mice. p < 0.0001 for vehicle vs Lac-Phe and Lac-Phe vs Lac-Phe+Tol. (c) Representative current-clamp traces showing effects of vehicle, Lac-Phe (5 μM), or Lac-Phe (5 μM) plus tolbutamide (100 μM) on AgRP neurons in the ARH of female mice. (d-e) Temporal changes in the resting membrane potential (d) and firing frequency (e) of AgRP neurons in the ARH in response to various treatments as described in (c). N = 6 neurons from 3 different female mice. p < 0.0001 for vehicle vs Lac-Phe and Lac-Phe vs Lac-Phe+Tol in both d and e. Data are mean ± SEM with individual data points in (b) and mean ± SEM in (d, e). Two-way ANOVA analysis followed by post hoc Sidak tests was used in (b, d, e). *, p < 0.05; **, p < 0.01; ****, p < 0.0001.
Extended Data Fig. 10 Validation of AAV-sgKcnj11.
(a) A schematic illustration for stereotaxic injections of AAV-spCas9 and AAV-sgKcnj11-FLEX-mCherry (ARHΔKcnj11), or AAV-GFP (control) into the ARH of male WT mice. (b) Three-hour food intake in HFD-fed male control (N = 8) or ARHΔKcnj11 (N = 9) mice (4 months of age) receiving i.p. injection of vehicle or Lac-Phe (50 mg/kg, i.p.). p < 0.0001 for vehicle vs Lac-Phe in control mice, p = 0.0039 for vehicle vs Lac-Phe in ARHΔKcnj11 mice, #### represents p < 0.0001 for Lac-Phe treated control vs ARHΔKcnj11 mice. (c) A representative microscopic image showing mCherry in the ARH of AgRP-IRES-Cre/Cas9fl/+ mice following AAV-sgKcnj11-FLEX-mCherry injection. N = 8 biological replicates. (d) Body weight gain in control and AgRPΔKcnj11 mice after HFD feeding. N = 8 mice in each group. p = 0.0069. (e) A representative current-clamp trace showing effects of diazoxide on AgRP neurons in the ARH of control mice. (f-g) Changes in the resting membrane potential (f) and firing frequency (g) of AgRP neurons in control mice following diazoxide treatment. N = 5 neurons. p = 0.0133 for f, p = 0.0138 for g. (h) A representative current-clamp trace showing effects of diazoxide on AgRP neurons in the ARH of AgRPΔKcnj11 mice. (i-j) Changes in the resting membrane potential (i) and firing frequency (j) of AgRP neurons in AgRPΔKcnj11 mice following diazoxide treatment. N = 6 neurons. (k) Representative voltage-clamp traces for KATP currents in AgRP neurons of control and AgRPΔKcnj11 mice in response to diazoxide. (l) Quantifications of KATP currents in AgRP neurons of control (N = 5) and AgRPΔKcnj11 (N = 6) mice. p = 0.00045. (m) Three-hour food intake in HFD-fed male control or ARHΔKcnj11 mice (4 months of age) receiving i.p. injection of vehicle or lorcaserin (9 mg/kg, i.p.). N = 5 mice in each group. p = 0.0136 and 0.0015 for vehicle vs lorcaserin in control and ARHΔKcnj11 mice, respectively. (n) Three-hour food intake in HFD-fed male control or AgRPΔKcnj11 mice (4 months of age) receiving i.p. injection of vehicle or lorcaserin (9 mg/kg, i.p.). N = 8 mice in each group. p < 0.0001 for vehicle vs lorcaserin in control and AgRPΔKcnj11 mice. Data are mean ± SEM with individual data points in (b, l-n) and mean ± SEM in (d) or individual data points in (f-g, i-j). Two-sided unpaired t-tests were used in (l); two-sided paired t-tests were used in (f-g, i-j); two-way ANOVA analysis was used in (b, d, m-n). *, p < 0.05; **, p < 0.01; ***, p < 0.001; ****, p < 0.0001.
Supplementary information
Source data
Source Data Figs. 1–4, Extended Data Figs. 1–10 (download XLSX )
Statistical source data for Figs. 1–4, Extended Data Figs. 1–10.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Liu, H., Li, V.L., Liu, Q. et al. Lac-Phe induces hypophagia by inhibiting AgRP neurons in mice. Nat Metab 7, 2004–2017 (2025). https://doi.org/10.1038/s42255-025-01377-9
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s42255-025-01377-9
This article is cited by
-
Lac-Phe: An Emerging Appetite-Regulating Exerkine with Health and Therapeutic Benefits
Current Obesity Reports (2026)
