
Abstract
The mitochondrial proteome is remodelled to meet metabolic demands, but how metabolic cues regulate mitochondrial protein turnover remains unclear. Here we identify a conserved, nutrient-responsive mechanism in which the amino acid leucine suppresses ubiquitin-dependent degradation of outer mitochondrial membrane (OMM) proteins, stabilizing key components of the protein import machinery and expanding the mitochondrial proteome to enhance metabolic respiration. Leucine inhibits the amino acid sensor GCN2, which selectively reduces the E3 ubiquitin ligase cofactor SEL1L at mitochondria. Depletion of SEL1L phenocopies the effect of leucine, elevating OMM protein abundance and mitochondrial respiration. Disease-associated defects in leucine catabolism and OMM protein turnover impair fertility in Caenorhabditis elegans and render human lung cancer cells resistant to inhibition of mitochondrial protein import. These findings define a leucine–GCN2–SEL1L axis that links nutrient sensing to mitochondrial proteostasis, with implications for metabolic disorders and cancer.
Similar content being viewed by others
Main
Mitochondrial proteostasis ensures that mitochondria can adjust their function to meet changing demands. Multiple quality control pathways, including mitochondria-associated degradation1,2,3,4,5,6, mitochondrial protein translocation-associated degradation7, mitochondria-derived vesicles8,9,10, mitochondria-derived compartments11,12 and mitophagy13,14 support this dynamic regulation. The mitochondrial proteome is highly adaptable; for example, the total mitochondrial protein mass in yeast more than doubles under respiratory versus fermentative growth conditions15. However, the specific nutrient signals and mechanisms that control this metabolic reshaping of the mitochondrial proteome remain unknown.
Communication between the mitochondria and the cytosol is critical for cellular adaptation to metabolic changes. The outer mitochondrial membrane (OMM), located at the interface between the cytosol and mitochondria, plays a key role in this crosstalk. OMM proteins coordinate protein import16,17, metabolite transport18,19,20, mitochondrial fusion and fission21 and interactions with other organelles22. Altered OMM proteostasis could thus coordinate changes in the cellular environment with mitochondrial activity. Nevertheless, whether and how nutrient signals impact OMM proteostasis for cellular adaptation remains largely unexplored.
Degradation of many OMM proteins is regulated by the ubiquitin-proteasome system (UPS)3,4,5,6,23,24,25,26,27,28,29,30,31, we therefore explored the role of ubiquitin-dependent regulation of OMM proteostasis in metabolic adaptation. To identify metabolic control of OMM protein stability, we conducted a genetic screen in Caenorhabditis elegans using a newly developed green fluorescent protein (GFP)-based reporter system that monitors UPS-mediated degradation of OMM-localized substrates. This approach unexpectedly revealed amino acid metabolism, particularly the branched-chain amino acid (BCAA) leucine (Leu), as an essential modulator of OMM protein degradation that tunes respiratory activity, dependent on the conserved amino acid sensor GCN-2. The HRD1 E3 ubiquitin ligase cofactor SEL1L is downregulated by Leu specifically at mitochondria, which leads to the stabilization of OMM substrates and enhancement of mitochondrial respiration. A disease-associated mutation in BCAT2, a key enzyme in Leu catabolism, stabilizes OMM proteins and impairs fertility under stress in C. elegans. Moreover, human lung cancer cells with elevated intracellular BCAA levels exhibit reduced OMM ubiquitylation and increased resistance to mitochondrial import inhibition. Together, our results identify a conserved mechanism by which Leu regulates OMM proteostasis and mitochondrial function, linking amino acid availability to mitochondrial remodelling and organismal health.
Results
A model substrate for monitoring OMM protein degradation
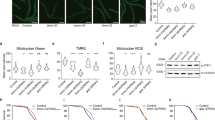
To monitor UPS-mediated degradation of OMM proteins, we developed a GFP-based ubiquitin fusion degradation (UFD) reporter assay in the multicellular organism C. elegans (Fig. 1a). UFD substrates, in which a protein is fused to a non-cleavable ubiquitin, are short-lived due to their targeting to the UPS32,33. Briefly, we fused non-cleavable ubiquitin–GFP (UbV–GFP) to the transmembrane domain of the OMM protein mitochondrial fission protein 1 (FIS-1) (Fig. 1a). The resulting OMM-anchored UFD substrate, termed mitoUFD is ubiquitously expressed in all C. elegans tissues, colocalizes with the OMM protein translocase of outer mitochondrial membrane 20 (TOMM-20)34 and surrounds the mitochondrial matrix (Fig. 1b). CytoUFD, a UbV–GFP fusion protein lacking the FIS-1-derived transmembrane domain, by contrast, did not colocalize with TOMM-20 (Fig. 1b). Next, we assessed mitoUFD turnover, using mitoGFP as a negative control, which is the same construct but lacking the ubiquitin as the degradation signal. Treatment of worms with the proteasome inhibitor bortezomib (BTZ) increased the abundance of mitoUFD, but not mitoGFP, at the mitochondrial surface (Fig. 1c,d and Extended Data Fig. 1a,b). In addition, overexpression of the proteasome subunit rpn-6.1 significantly reduced mitoUFD levels (Extended Data Fig. 1c,d). Furthermore, the cycloheximide (CHX) chase assay demonstrated fast turnover of mitoUFD while the levels of the control mitoGFP remained stable (Fig. 1e,f). Taken together, we conclude that mitoUFD is targeted for degradation by the UPS.

a, The mitoUFD model substrate is located at the OMM and a substrate for the UPS. IMM, inner mitochondrial membrane; Ub, ubiquitin. b, Confocal images showing that mitoUFD colocalizes with a previously established OMM marker TOMM-20::mKate2 (top row), surrounding the mitoTracker-stained mitochondrial matrix (middle row), and cytoUFD does not colocalize with TOMM-20::mKate2 (bottom row). Scale bar, 10 µm. c, Confocal images showing that BTZ treatment at 5 µM or 10 µM concentration stabilizes mitoUFD at the OMM. Scale bar, 10 µm. d, Western blot analysis showing that BTZ treatment stabilizes mitoUFD but not mitoGFP. e, Western blot showing chase assay of mitoUFD (degraded) versus mitoGFP (stable). f, Quantification of GFP levels in worms by fluorescence imaging over a 9 h CHX-chase assay. Mean ± s.e.m., n = 5 independent biological replicates. Two-way ANOVA; the P value indicates the effect of substrates (mitoUFD versus mitoGFP). g, A volcano plot showing enriched binding partners of mitoUFD compared with mitoGFP, which includes ubiquitin associated/related proteins and proteasome subunits. h, Mitochondrial enrichment followed by pulldown of polyubiquitylated proteins by TUBE agarose in L4440 control and cdc-48 RNAi worms; western blot (WB) against GFP to show polyubiquitylated mitoUFD substrate. Input: isolated and lysed mitochondria before TUBE IP. The same number of worms was used for both conditions.
In yeast, the AAA+ ATPase Cdc48 (p97 in vertebrates)35,36 and its cofactors extract client OMM proteins for proteasomal degradation1,3 (Extended Data Fig. 1e). By pulldown of the mitoUFD substrate, we identified CDC-48 as its binding partner, as well as some ubiquitin associated/related proteins and subunits of the proteasome (Fig. 1g). RNA interference knockdown of cdc-48/CDC48 or its cofactors ufd-3/DOA1, npl-4/NPL4 and ufd-1/UFD1 increased mitoUFD but not mitoGFP levels at the mitochondrial surface (Extended Data Fig. 1f–h). In addition, cdc-48 RNAi knockdown animals accumulated ubiquitylated forms of mitoUFD (Fig. 1h). These results suggest that CDC-48-dependent extraction of mitoUFD from the OMM is required for its proteasomal degradation.
Amino acid metabolism affects OMM protein degradation
To investigate how mitochondrial homeostasis affects OMM protein degradation, we used RNAi to knockdown 38 nuclear-encoded mitochondrial proteins that have mammalian orthologues and belong to different mito-pathways in the MitoCarta 3.0 datasets37, including mitochondrial central dogma; protein import, sorting and homeostasis; small-molecule transport; signalling; mitochondrial dynamics; surveillance and oxidative phosphorylation (Extended Data Fig. 2a,b and Supplementary Table 1). In most cases, depletion of these mitochondrial proteins (24/38) increased mitoUFD levels but did not affect, or even decreased, cytoUFD levels (Extended Data Fig. 2c). By contrast, depletion of genes encoding subunits of the respiratory chain complexes had a weaker effect on stabilizing mitoUFD compared to cytoUFD (Extended Data Fig. 2c).
We noticed that knockdown of SLC-25A42 led to a consistently strong elevation of mitoUFD (Extended Data Fig. 2c). SLC-25A42 is a mitochondrial transporter for coenzyme A (CoA)38, which forms acetyl-CoA in the mitochondria. Acetyl-CoA is involved in many biochemical reactions, most notably the tricarboxylic acid (TCA) cycle. To determine whether other genes involved in the TCA cycle affect mitoUFD, we downregulated eight key enzymes for TCA activity (Extended Data Fig. 2d). Indeed, knockdown of three (aco-2, idh-1 and sdhb-1) significantly increased mitoUFD levels (Extended Data Fig. 2e), suggesting a general role for the TCA cycle in regulating UPS-dependent mitoUFD degradation.
Disruption of the TCA cycle has a profound effect on amino acid metabolism39,40. TCA cycle intermediates serve as building blocks for the synthesis of various amino acids41,42,43, and amino acid catabolism produces intermediates such as acetyl-CoA that feed into the TCA cycle. Given this tight coupling, we hypothesized that disrupting amino acid metabolism might affect mitoUFD levels. To test this, we performed a genetic screen of 135 genes that encode enzymes involved in amino acid metabolism44 (Supplementary Table 2). Knockdown of 67/135 enzymes involved in amino acid metabolism increased mitoUFD (Fig. 2a and Extended Data Fig. 3a), suggesting that amino acid metabolism has a broad impact on OMM protein degradation. By contrast, knockdown of genes involved in amino acid transport (aat-1, aat-2, aat-3 and F13G3.7), glucose sensing (egl-30 and gpa-4)45,46 and metabolite transport (C16C10.1, slc-25A21 and slc-25A29)12,38 had little or no effect on mitoUFD (Extended Data Fig. 3b). Of these 67 genes, knockdown of 16 led to a >20% increase in mitoUFD levels (Fig. 2a and Extended Data Fig. 3a). These top 16 genes belong to different metabolic pathways involving 14 amino acids (Extended Data Fig. 3c).

a, RNAi knockdown of amino acid metabolic genes has a broad effect on mitoUFD degradation. Fifty per cent of the amino acid metabolic genes stabilize mitoUFD by more than 10% (GFP intensity of 50–250 animals for each gene was measured). b, Changes in mitoUFD and cytoUFD levels upon RNAi knockdown of genes involved in amino acid sensing. Mean ± s.e.m., n = 4 independent biological replicates with a total of 24–32 animals. ImageJ quantification. Two-way ANOVA with Holm–Sidak correction for multiple comparisons with L4440 control within mitoUFD or cytoUFD group. c, Double knockdown of hoe-1 and gcn-2 does not show an additive effect on mitoUFD degradation. Mean ± s.e.m., n = 4 independent biological replicates with a total of 24–32 animals. ImageJ quantification. Unpaired two-tailed t-test between double knockdown and single-knockdown control. d, BCAA catabolic pathway and the key enzymes involved. The five candidates indicated with arrowhead are among the top hits from the targeted genetic screen in a. e, Changes in mitoUFD and cytoUFD levels upon RNAi knockdown of the five genes involved in BCAA catabolism in the top hits. Quantification by worm sorter. Mean ± s.e.m., n = 3 independent biological replicates with 50–250 animals in each replicate. One-way ANOVA with Fisher’s least significant difference (LSD) test for multiple comparisons was performed using L4440 as the control for both mitoUFD and cytoUFD. While the knockdown of mccc-1 and ech-1.2 did not result in statistically significant changes in cytoUFD (P > 0.05), the average percentage changes were greater than 10%. f, After Leu, isoleucine or valine supplementation for 3 h, mitoUFD and cytoUFD levels were measured. Mean ± s.e.m., n = 4 independent biological replicates (with a total of 24–32 animals). ImageJ quantification. Two-way ANOVA with Holm–Sidak correction for multiple comparisons with L4440 control within the mitoUFD or cytoUFD group. g, Leu supplementation stabilizes mitoUFD at 20 mM and 50 mM. mitoGFP was used for control. Mean ± s.e.m., n = 6 independent biological replicates for mitoUFD (with a total of 36–48 animals) and n = 8 independent biological replicates for cytoUFD (with a total of 48–64 animals). ImageJ quantification. Two-way ANOVA with Holm–Sidak correction for multiple comparisons with 0 mM Leu control. h, RNAi knockdown of gcn-2 but not let-363 followed by 3 h Leu supplementation abolished Leu-induced increases in mitoUFD levels. Mean ± s.e.m., n = 4 independent biological replicates with a total of 24–32 animals. ImageJ quantification. One-way ANOVA with Holm–Sidak correction for multiple comparisons with L4440 control. i, Western blot showing polyubiquitylated mitoUFD substrate expressed in HEK293 cells. Cells treated with BTZ at 0.2 or 1 µM showed stabilization of mitoUFD compared with control treatment. j, Confocal imaging of HEK293 cells expressing mitoUFD after treatment with 0, 1 or 3 mM Leu for 3 h. Scale bar, 20 µm. k, CHX chase assay showing the increased stability of human mitoUFD upon 1 mM Leu treatment. Mean ± s.e.m., n = 4 independent biological replicates. Two-way ANOVA; the P value indicates Leu treatment effect. NS, not significant.
Given this widespread influence, we hypothesized that amino acid sensor(s) might link amino acid availability to mitoUFD degradation. We focused on GCN-2, a cytosolic kinase that is activated by uncharged tRNA in response to amino acid imbalance47,48,49,50. Knockdown of gcn-2 resulted in a strong, selective increase in mitoUFD (Fig. 2b and Extended Data Fig. 4a). Knockdown of hoe-1, which encodes the tRNA 3′ processing enzyme whose loss is known to impair tRNA maturation, also increased mitoUFD levels (Fig. 2b), consistent with reduced activation of GCN-2 under tRNA deficiency50. Double knockdown of gcn-2 and hoe-1 increased mitoUFD levels similarly to knockdown of gcn-2 alone (Fig. 2c), suggesting that they act in the same pathway. Importantly, control mitoGFP levels were not affected by knockdown of gcn-2 (Extended Data Fig. 3d), indicating that GCN-2 is specifically required for UPS-mediated turnover. Since amino acids are well known to activate mTORC1 signalling51, we also examined the C. elegans orthologue of mTOR let-363 and the mTORC1 downstream effector rsks-1/S6K. Knockdown of let-363 did not affect mitoUFD and knockdown of rsks-1 only slightly increased mitoUFD levels (Fig. 2b), suggesting that the regulation of OMM protein degradation is independent of mTORC1 signalling. These data reveal that GCN-2 links amino acid availability to changes in protein degradation at the OMM.
Leu selectively inhibits OMM protein degradation
We next asked which amino acid(s) regulate mitoUFD levels. Among the top hits in our screen were five enzymes involved in the catabolism of the BCAAs Leu, isoleucine and valine (bcat-1, dld-1, mccc-1, ech-1.2 and T09B4.8; Fig. 2d and Extended Data Fig. 3c). We validated the five genes and compared their impact on the degradation of mitoUFD and cytoUFD. Knockdown of bcat-1, which encodes the first enzyme in the BCAA catabolic pathway, increased mitoUFD but not cytoUFD, whereas knockdown of the other four downstream enzymes increased both mitoUFD and cytoUFD levels (Fig. 2e and Extended Data Fig. 4a). These data recapitulate the effects of GCN-2 and suggest a specific role for BCAT-1 in the regulation of mitoUFD.
As the loss of bcat-1 increases cellular levels of free BCAAs52,53, we hypothesized that elevated BCAA concentrations would mimic depletion of bcat-1. To test this, we treated the worms with high concentrations of Leu, isoleucine or valine. We found that 3 h of treatment with Leu, but not isoleucine or valine, significantly increase mitoUFD levels without affecting cytoUFD (Fig. 2f and Extended Data Fig. 4b), phenocopying gcn-2 and bcat-1 knockdowns. mitoGFP levels were not affected (Fig. 2g), indicating that Leu does not broadly alter OMM protein levels but specifically regulates UPS-dependent OMM protein degradation. Notably, Leu supplementation failed to elevate mitoUFD levels when GCN-2 was depleted, whereas Leu was still able to elevate mitoUFD levels when let-363 was knocked down (Fig. 2h), suggesting that high levels of Leu reduce proteolytic activity at the OMM via GCN-2-mediated amino acid sensing independent of mTOR.
To determine whether the Leu-mediated regulation of mitoUFD is evolutionarily conserved, we expressed mitoUFD in human HEK293 cells. The mitoUFD substrate was ubiquitylated and targeted for proteasomal degradation in HEK293 cells (Fig. 2i and Extended Data Fig. 4c,d) and it displayed increased stability in the presence of excess Leu (Fig. 2j,k). These data suggest that Leu-dependent regulation of OMM protein degradation is conserved in worms and human cells.
Leu decreases ubiquitylation of OMM proteins via SEL-1–SEL-11
We next asked how Leu treatment increases the stability of OMM proteins. We did not observe any significant differences in the activity of the 26S proteasome between HEK293 cells with and without Leu treatment (Extended Data Fig. 5a,b). By contrast, global ubiquitylation of OMM proteins was significantly reduced by Leu supplementation, whereas whole-cell ubiquitylation was not affected (Fig. 3a and Extended Data Fig. 5c). Furthermore, cells treated with a GCN-2 inhibitor54 showed a substantial reduction in OMM protein ubiquitylation, comparable to that achieved with Leu supplementation, and these effects were not additive (Fig. 3b). These results suggest that excess Leu specifically reduces OMM but not cytosolic protein ubiquitylation via GCN-2 inhibition.

a, Ubiquitylation of whole cell proteins and OMM proteins of cells cultured in medium supplemented with 0 mM or 1 mM Leu and/or 0 µM or 1 µM BTZ. For each sample, 10 µg proteins were loaded. Ubiquitylation levels were normalized to the total OMM protein amount. Mean ± s.e.m., n = 4 independent biological replicates. Unpaired two-tailed t-test. b, Ubiquitylation of whole-cell proteins and OMM proteins of cells cultured in medium supplemented with 0 mM or 1 mM Leu and/or 0 µM or 10 µM GCN-2 inhibitor for 3 h. For each sample, 10 µg proteins were loaded. Ubiquitylation levels were normalized to the total OMM protein amount. Mean ± s.e.m., n = 3 independent biological replicates. Unpaired two-tailed t-test. c, A volcano plot showing the proteomic analysis of Leu (20 mM, 3 h treatment) on enriched mitochondria (mito). P values were determined by an unpaired two-tailed t-test. E3 ligases/cofactors are labelled in red. SEL-1 was the only significantly downregulated (−log10P >1.3) E3 ligase/cofactor in enriched mitochondria upon Leu treatment. d, Western blot quantification of SEL-1 in enriched mitochondria and whole worms. Animals were treated with and without 20 mM Leu supplementation for 3 h. Unpaired two-tailed t-test. Mean ± s.e.m., n = 5 independent biological replicates. e, CHX chase assay showing the increased stability of mitoUFD substrate in worms upon RNAi knockdown of sel-1 and sel-11. Mean ± s.e.m., n = 3 independent biological replicates. f, Confocal imaging showing mitoUFD stabilization at the OMM upon RNAi knockdown of sel-1 or sel-11. Stabilized mitoUFD is colocalized with TOMM-20. g, ImageJ quantification of mitoUFD levels upon RNAi knockdown of sel-1 or sel-11. Mean ± s.e.m., n = 4 independent biological replicates with 24–32 animals in each replicate. One-way ANOVA with Holm–Sidak correction for multiple comparisons with L4440 control. h, Western blot quantification of SEL1L and HRD1 protein level with and without 3 h Leu supplementation in the whole-cell, mitochondrial (P10) and microsome (P100) fraction. Mean ± s.e.m., n = 7 independent biological replicates. Unpaired two-tailed t-test.
To determine how Leu reduces the ubiquitylation of OMM proteins, we performed proteomic analysis to identify ubiquitin ligases whose levels changed in response to Leu supplementation. We treated worms with 20 mM Leu or with dH2O. After 3 h of treatment, whole-worm lysates and mitochondria-enriched fractions were analysed by mass spectrometry. Interestingly, we found that SEL-1, the worm orthologue of SEL1L, which is known to form an E3 ubiquitin ligase complex together with HRD1 (SEL-11 in C. elegans) at the endoplasmic reticulum membrane55,56, was significantly reduced upon Leu treatment in the mitochondrial fraction, and this change was not observed in the whole worm (Fig. 3c and Extended Data Fig. 6a). HRDL-1 and RNF-126 were also downregulated in the mitochondrial fraction, but their changes were not statistically significant (−log10P <1.3) (Fig. 3c). Therefore, we focused on SEL-1. We confirmed that the decrease in SEL-1 abundance was specific to the mitochondrial fraction by western blot (Fig. 3d). Moreover, the levels of the endoplasmic reticulum-associated degradation (ERAD) substrate CPL-1*, whose degradation is mediated by SEL-1 and SEL-11 (ref. 57), were not stabilized by Leu supplementation (Extended Data Fig. 6b,c).
SEL-1 is membrane bound in worms, and cofractionates with the OMM-anchored mitoUFD (Extended Data Fig. 6d). SEL-1 and SEL-11 exhibit enriched binding to mitoUFD compared with mitoGFP by more than two fold (Extended Data Fig. 6e), suggesting that the SEL-1–SEL-11 E3 ligase complex directly interacts with and ubiquitylates OMM proteins. Immunogold analysis further supported the localization of SEL-1 to both the endoplasmic reticulum and OMM (Extended Data Fig. 6g,h). We performed RNAi knockdown of sel-1 and sel-11 and indeed observed an increased stability of the mitoUFD substrate at the OMM (Fig. 3e–g and Extended Data Fig. 6f), suggesting that the reduced abundance of SEL-1 due to high levels of Leu limits ubiquitin-dependent degradation of OMM proteins. In addition, in HEK293 cells, Leu supplementation reduced the level of mitochondrial-associated SEL1L, whereas the level of SEL1L in the endoplasmic reticulum-derived microsomal fraction remained unchanged (Fig. 3h), suggesting a conserved regulation of SEL-1/SEL1L by Leu at the OMM in worms and human cells. Together, our results suggest that high Leu concentrations reduce SEL-1–SEL-11-dependent degradation of OMM proteins.
Leu supplementation triggers OMM proteome expansion
To determine whether Leu regulates endogenous OMM proteins, we re-analysed our proteomics data by filtering mitochondrial proteins using MitoCarta 3.0 and OMM annotations (Fig. 4a). We observed an overall increase in the mitochondrial and OMM proteome, but not in the total worm proteome upon Leu supplementation (Extended Data Fig. 7a–d). To evaluate whether specific protein classes were regulated by Leu supplementation, we performed Gene Ontology (GO) enrichment analysis and found that mitochondrial transporters, protein processing and organization, TCA cycle and respiratory chain activity were highly enriched for upregulated proteins (Extended Data Fig. 7e).

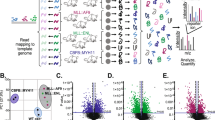
a, The experimental paradigm of proteomic profiling of whole worms and mitochondria of worms supplemented with dH2O, 20 mM Leu, CHX and Leu + CHX for 3 h. Four independent biological replicates were analysed for each condition. b, The percentage of proteins increased or decreased in the whole-worm proteome, MitoCarta 3.0 proteins and OMM proteins for each comparison indicated. c, The log2 fold change of the whole-worm proteome, MitoCarta 3.0 proteins and OMM proteins comparing Leu + CHX with CHX treatment. Two-tailed Mann–Whitney U test between whole-worm proteins and other protein groups. d, Volcano plot showing changes of OMM proteins upon Leu + CHX treatment compared with CHX treatment. P values were determined by an unpaired two-tailed t-test. OMM proteins show the tendency to be more abundant in Leu-treated worms in the presence of CHX. e, Volcano plot showing changes of whole worm and MitoCarta 3.0 proteins upon Leu + CHX treatment compared with CHX treatment. P values were determined by an unpaired two-tailed t-test. Mitochondrial proteins show the tendency to be more abundant in Leu-treated worms in the presence of CHX. f, The experimental paradigm of proteomic profiling of HEK293 whole cells and mitochondria of cells supplemented with dH2O, Leu, CHX and Leu + CHX for 3 h. Four independent biological replicates were analysed for each condition. g, The percentage of proteins increased or decreased in the whole cell proteome, MitoCarta 3.0 proteins and OMM proteins in each comparison indicated. h, The log2 fold change of the whole cell proteome, MitoCarta 3.0 proteins and OMM proteins comparing Leu + CHX with CHX treatment. Two-tailed Mann–Whitney U test between whole cell proteins and other protein groups. i, A volcano plot showing changes of OMM proteins upon Leu + CHX treatment compared with CHX treatment. P values were determined by an unpaired two-tailed t-test. OMM proteins show the tendency to be more abundant in Leu-treated cells in the presence of CHX. j, A volcano plot showing the changes in whole cell and MitoCarta 3.0 proteins upon Leu + CHX treatment compared with CHX treatment. P values were determined by an unpaired two-tailed t-test. Mitochondrial proteins show the tendency to be more abundant in Leu-treated cells in the presence of CHX. k, Enriched GO terms (P < 0.001 and FDR < 0.2 cut-off) for upregulated proteins from the worm mitochondrial proteome in the Leu + CHX condition compared with the CHX condition. l, Enriched GO terms (P < 0.001 and FDR <0.2 cut-off) for upregulated proteins from the HEK293 cell mitochondrial proteome in the Leu + CHX condition compared with the CHX condition. A full list of significantly enriched GO terms is presented in Supplementary Table 3 for worm and HEK293 cells.
To exclude a possible effect of Leu on protein translation, we repeated the proteomic analysis in the presence of CHX to block cytosolic translation (Fig. 4a). CHX treatment did not eliminate the overall increase in OMM proteins upon Leu treatment (Fig. 4b–d and Extended Data Fig. 7f), supporting our hypothesis that the increase in OMM proteins is the result of reduced post-translational protein degradation. We found that nine OMM proteins in worms were consistently upregulated in response to Leu treatment in two independent proteomic analyses and were also upregulated by Leu in the presence of CHX (Fig. 4d and Extended Data Fig. 7b,f). The nine OMM proteins are DNJ-9 (DNAJC11, regulation of protein import and sorting), FKB-6 (FKBP8, regulation of apoptosis), GOP-3 (SAMM50, assembly of TOMM-40 into the TOM complex), MTCH-1 (MTCH1/2, protein insertase that mediates insertion of transmembrane proteins into the OMM), PGAM5 (PGAM5, regulation of mitophagy and mitochondrial dynamics), TOMM-40 (TOMM-40, subunit of the TOM complex, import of protein precursors into mitochondria), VDAC-1 (VDAC1/2/3, the channel at the OMM allows the transport of calcium and small molecules), KMO-1 (KMO, mitochondrial dynamics) and FZO-1 (MFN1/2, mitochondrial fusion). Interestingly, four of them (TOMM-40, GOP-3, MTCH-1 and DNJ-9) are involved in mitochondrial protein import and assembly, suggesting that Leu treatment increases mitochondrial protein import capacity. Indeed, there was a consistent upregulation of the total mitochondrial proteome upon Leu treatment in two independent proteomic analyses (Extended Data Fig. 7c,g), which was maintained in the presence of CHX (Fig. 4b,c,e). CHX treatment alone depleted mitochondrial proteins (Fig. 4b and Extended Data Fig. 7h). As CHX specifically reduces translation in the cytosol while leaving mitochondrial translation unaffected58, the reduced mitochondrial protein content was probably due to reduced import as a compensatory response to arrested cytosolic protein synthesis, in agreement with a previous report59.
To determine whether this response is conserved, we performed the same experiment with HEK293 cells. We observed a striking increase in the abundance of the OMM proteome in the presence of Leu regardless of CHX treatment (Fig. 4f–j and Extended Data Fig. 7i–k), including seven orthologues of the nine C. elegans OMM proteins regulated by Leu. Again, several upregulated proteins, such as SAMM50, DNAJC11, TOMM-40 and MTCH1/2, are involved in protein import and assembly (Fig. 4i). We confirmed the increased protein stability of TOMM-40 upon Leu supplementation by western blot (Extended Data Fig. 8a,b). The conserved increase in these proteins along with the mitochondrial proteome upon Leu supplementation suggests a fundamental role for Leu in rewiring the mitochondrial proteome by modulating the import machinery. GO term analysis revealed a consistent enrichment of upregulated mitochondrial proteins involved in mitochondrial transport, amino acid metabolism, TCA cycle and respiratory chain activity in both worms and HEK293 cells (Fig. 4k,l and Supplementary Table 3), suggesting an overall increase in mitochondrial metabolic activity in response to excess Leu.
Together, our results suggest that Leu expands the OMM proteome by reducing the degradation of endogenous OMM proteins, including the subunit of the TOM complex TOMM-40, which allows the import of protein precursors into mitochondria, thereby remodelling the mitochondrial proteome involved in metabolism and respiration.
Leu-driven OMM remodelling boosts mitochondrial respiration
We hypothesized that the reduced OMM protein degradation and increased abundance of the mitochondrial proteome in response to Leu treatment would improve mitochondrial activity. Indeed, 3 h of Leu treatment resulted in increased mitochondrial respiration in both worms and human cells (Fig. 5b and Extended Data Fig. 9a,b). Notably, although treating HEK293 cells with other essential amino acids resulted in a slight increase in mitochondrial respiration, Leu had the highest impact (Extended Data Fig. 9a,b).

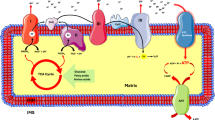
a, cdc-48.1 overexpression (OE) reduces mitoUFD levels. Mean ± s.e.m., n = 4 independent biological replicates. Unpaired two-tailed t-test. b, Seahorse OCR measurement for WT and cdc-48.1 OE animals with/without 50 mM Leu supplementation. Two-way ANOVA with Fisher’s LSD test for multiple comparisons. Mean ± s.e.m., n = 4 independent biological replicates. c, The percentage of proteins increased or decreased in whole-worm proteome, MitoCarta 3.0 proteins and OMM proteins in Leu-treated cdc-48.1 OE animals. d, A volcano plot showing changes of OMM proteins upon Leu treatment. P values were determined by an unpaired two-tailed t-test. e, A volcano plot showing changes of whole-worm and MitoCarta 3.0 proteins upon Leu treatment. P values were determined by an unpaired two-tailed t-test. f, The log2 fold change of whole-worm proteome, MitoCarta 3.0 proteins and OMM proteins upon Leu treatment. Two-tailed Mann–Whitney U test between whole-worm proteins and other protein groups. g, gcn-2 OE reduces mitoUFD levels. Mean ± s.e.m., n = 4 independent biological replicates. Unpaired two-tailed t-test. h, Seahorse OCR measurement for control SCR OE and gcn-2 OE animals with or without 50 mM Leu supplementation. Two-way ANOVA with Fisher’s LSD test for multiple comparisons. Mean ± sem, n = 4 independent biological replicates. Statistical analysis in Extended Data Fig. 9e. i, Seahorse OCR measurement for control L4440 RNAi and tomm-40 RNAi animals with or without 50 mM Leu supplementation. Two-way ANOVA with Fisher’s LSD test for multiple comparisons. Mean ± s.e.m., n = 4 independent biological replicates. Statistical analysis in Extended Data Fig. 9f. j, Seahorse OCR measurement for control L4440 RNAi and sel-1, sel-11 RNAi animals. One-way ANOVA with Fisher’s LSD test for multiple comparisons. Mean ± s.e.m., n = 4 independent biological replicates. Statistical analysis in Extended Data Fig. 9i. k, bcat-1(E279K)/+ worms showed higher mitoUFD levels compared with WT animals. Mean ± s.e.m., n = 3 independent biological replicates with a total of 18–24 animals. ImageJ quantification. l, Brood size of WT and bcat-1(E279K)/+ animals with gcn-2 RNAi knockdown from L4 stage compared with L4440 control knockdown. One-way ANOVA with Fisher’s LSD test for multiple comparisons. m, Brood size of animals with gcn-2 OE from L4 stage. Unpaired two-tailed t-test. n, BCAA levels in H2030, H1437 and H1666. Unpaired two-tailed t test. Mean ± s.e.m., n = 3 independent biological replicates with 3 technical replicates each biological replicate. o, Ubiquitylation of OMM proteins in H2030, H1437 and H1666. Mean ± s.e.m., n = 3 independent biological replicates. Ubiquitylation levels were normalized to total OMM protein amount. Unpaired two-tailed t-test. p–r, The cell viability of H2030 (p), H1437 (q) and H1666 (r) over 4 days with 0 µM, 4 µM and 10 µM MB-12 treatment. Two-way repeated ANOVA; the P value indicates the MB-12 treatment effect. Mean ± s.e.m., n = 3 independent biological replicates with 3 technical replicates each day each condition. s, A model showing the regulation of OMM protein degradation by the Leu–GCN-2–SEL1L axis. High levels of Leu inhibit GCN-2 and reduce SEL1L–HRD1-dependent degradation of OMM proteins, stabilizing key components of the protein import machinery such as TOMM40. This leads to mitochondrial proteome expansion for respiratory chain, TCA cycle, metabolism and metabolite transport, thereby enhancing mitochondrial respiratory capacity and cell viability.
To test whether the Leu-induced increase in mitochondrial respiration requires elevated OMM protein abundance, we examined worms overexpressing cdc-48.1 (ref. 60) (Extended Data Fig. 9c). Overexpression of cdc-48.1 reduced the levels of mitoUFD (Fig. 5a), consistent with its role in extraction and degradation of OMM proteins. Notably, the maximal respiratory rate was lower in worms overexpressing cdc-48.1 compared with controls, and this rate was unaffected by Leu supplementation in contrast to controls (Fig. 5b). Proteomic analysis indicated that cdc-48.1 overexpression prevented the increase in the OMM and mitochondrial proteome in response to Leu (Fig. 5c–f). Similarly, we found that overexpression of GCN-2 reduced mitoUFD levels (Fig. 5g and Extended Data Fig. 9d) and prevented the Leu-induced increase in maximal respiration (Fig. 5h and Extended Data Fig. 9e). To investigate whether mitochondrial import is required for the Leu-dependent increase in mitochondrial respiration, we examined the role of TOMM-40. Acute tomm-40 knockdown abolished the Leu-dependent increase in their maximal respiratory rate (Fig. 5i and Extended Data Fig. 9f). Moreover, Leu treatment failed to enhance mitochondrial respiration in HEK293 cells when mitochondrial import was inhibited by MitoBlock-12 (MB-12) treatment61,62 (Extended Data Fig. 9g,h). This supports the idea that the Leu-induced increase in mitochondrial respiration is mediated by the mitochondrial protein import machinery.
Importantly, RNAi knockdown of sel-1 or sel-11 resulted in enhanced mitochondrial respiration (Fig. 5j and Extended Data Fig. 9i), phenocopying Leu supplementation. This observation suggests that the Leu-dependent reduction of SEL-1 and the resulting increased stability of OMM proteins enhanced mitochondrial respiration. Taken together, these results suggest that Leu acts via stabilizing OMM proteins, facilitating mitochondrial proteome expansion through the import machinery, to increase mitochondrial respiratory capacity.
BCAA–OMM proteostasis axis influences fertility and cell growth
Patients with an E264K mutation in BCAT2 (the orthologue of C. elegans bcat-1) have reduced BCAT2 activity and elevated plasma levels of BCAAs63. To determine how this disease-associated mutation affects organismal physiology, we used CRISPR–Cas9 to generate a bcat-1(E279K) mutant in C. elegans, which corresponds to human BCAT2E264K (Extended Data Fig. 10a)63. Homozygous mutant worms were lethal, so we examined heterozygous bcat-1(E279K/+) worms. We observed higher levels of mitoUFD throughout bcat-1(E279K/+) compared with wild-type (WT) animals (Fig. 5k and Extended Data Fig. 10b), suggesting reduced UPS activity at the OMM. Despite this phenotype, bcat-1(E279K/+) animals produced a normal number of eggs, with no differences in brood size or hatch rate compared with WT animals (Fig. 5l and Extended Data Fig. 10c). Given our data that gcn-2 regulates UPS activity at the OMM, we asked whether disrupting this regulatory node affects fitness in the context of impaired BCAA metabolism. To test this, we knocked down gcn-2 in bcat-1(E279K/+) animals. Knockdown of gcn-2 alone did not impact brood size or egg hatch rate in WT animals, but in the bcat-1(E279K/+) background, it caused a significant reduction in brood size (Fig. 5l). This genetic interaction suggests that bcat-1(E279K/+) animals are sensitized to further disruption of mitochondrial proteostasis. Overexpression of gcn-2, which enhanced OMM protein degradation (Fig. 5g), reduced egg laying in WT animals, without affecting hatch rate (Fig. 5m and Extended Data Fig. 10d). These results suggest that an optimal level of BCAA metabolism and OMM protein degradation is linked to animal fertility.
Defects in BCAA metabolism are associated with tumour growth and progression64,65. We measured intracellular BCAA levels in three non-small cell lung cancer cell lines, H2030, H1437 and H1666, and found that H2030 and H1666 have higher intracellular BCAA levels compared with H1437 (Fig. 5n). The increased BCAA level in H2030 is probably due to a loss-of-function mutation in the BCAT2 gene (Extended Data Fig. 10e). We isolated OMM proteins from the three lung cancer cell lines and found that OMM protein ubiquitylation was significantly lower in H2030 and H1666 compared with H1437 (Fig. 5o). Interestingly, blocking mitochondrial import with MB-12 substantially inhibited the proliferation of H1437 and HEK293 cells, but only mildly affected the proliferation of H2030 and H1666 cells with increased MB-12 concentration (Fig. 5p–r and Extended Data Fig. 10f–j). Collectively, our results suggest that the BCAA-dependent regulation of OMM proteostasis is crucial for maintaining animal fertility and cell viability.
Discussion
Our work reveals a conserved, nutrient-responsive mechanism that regulates mitochondrial proteome homeostasis through targeted modulation of protein ubiquitylation at the OMM. Using a novel reporter system (mitoUFD), we demonstrate that Leu, one of the essential BCAAs, stabilizes OMM proteins by decreasing their ubiquitin-dependent degradation. This process occurs through a conserved pathway involving GCN2 in response to Leu supplementation. As a result, key components of the mitochondrial import machinery, such as TOMM40, accumulate, thereby enhancing protein import, expanding the mitochondrial proteome particularly for proteins involved in metabolism and respiration, and boosting respiratory activity. Specifically, the SEL-1–SEL-11/SEL1L–HRD1 E3 ubiquitin ligase complex is required for OMM protein degradation, and the SEL-1/SEL1L level is regulated by Leu acting at the OMM for adjusting mitochondrial proteostasis and respiration. Together, these findings connect nutrient sensing and mitochondrial protein turnover with organismal health.
Previous studies have suggested a crosstalk between amino acid metabolism and mitochondrial homeostasis. For example, when impairment occurs to lysosomal function, which is important to import and sequester amino acids to avoid amino acid toxicity, cells can generate mitochondrial-derived compartments to remove the mitochondrial protein import receptor Tom70 and SLC25A carriers in response to amino acid overload stress12. Amino acid sensing also controls mitochondrial fusion to regulate mitophagy and apoptosis66. Long-term Leu exposure, for example, 48 h treatment, increases mitochondrial biogenesis and respiration in C2C12 muscle cell culture by activating SIRT1–AMPK signalling67. Instead of the conventional role of Leu in anabolism, our data uncover a distinct mechanism that acute Leu supplementation leads to a selective reduction in the ubiquitylation and degradation of OMM proteins, thereby remodelling the mitochondrial proteome independently of translation. This rewiring is indispensable for increasing the abundance of the import machinery and elevating mitochondrial respiration, potentially preparing cells for the bioenergetic demands of high nutrient availability (Fig. 5s).
The identification of the SEL-1–SEL-11/SEL1L–HRD1 complex acting specifically at the OMM and its modulation by Leu availability adds an important new dimension to our understanding of mitochondrial proteostasis. Recent studies have shown that the SEL1L–HRD1 complex binds to OMM proteins such as SAMM50, PGAM5 and VDAC1 and might regulate the stability of MTCH2 (ref. 68), implying a role for endoplasmic reticulum-associated E3 ligases and cofactors in OMM protein degradation. Furthermore, SEL1L–HRD1 has been shown to regulate mitochondrial dynamics by modulating endoplasmic reticulum–mitochondria contacts in brown adipocytes, which is essential for metabolic adaptation to cold challenge69. Other ERAD components, such as Cdc48/p97 and Ubx2, have also been implicated in mitochondrial protein quality control1,2,3,7,70,71,72, and there is evidence that Ubx2 localizes to both the endoplasmic reticulum and OMM7. Although we find that SEL-1 is membrane bound and associated with mitochondria, it lacks an obvious mitochondrial targeting sequence. Therefore, further investigation is required to determine how this E3 ligase complex is targeted to the OMM, whether endoplasmic reticulum–mitochondria contact sites play a role in this process and how Leu specifically regulates mitochondria-associated SEL-1.
Leu serves as an important building block for protein synthesis, a nutrient signal and an energy source. Altered BCAA/Leu metabolism and mitochondrial changes have been implicated in various diseases. For example, lifelong inhibition of BCAA metabolism leads to reproductive ageing and is associated with mitochondrial dysfunction in C. elegans53. Leu is also a key factor driving tumour growth of aggressive rhabdomyosarcoma via modulation of mitochondrial metabolism and oxidative phosphorylation65. Our work suggests that BCAA metabolism sustains degradation of OMM proteins and tunes mitochondrial activity, which is associated with fertility when encountering stress. By analysing lung cancer cells with different intracellular BCAA levels, we show that BCAAs are directly related to OMM proteostasis and cellular resistance to mitochondrial import inhibition, which may be an important aspect to consider for cancer treatment.
In conclusion, our study provides evidence that cytosolic nutrient status directly modulates OMM proteostasis as a rapid response to adjust mitochondrial activity. This reshaping of the OMM proteome enhances mitochondrial import and respiratory capacity, linking nutrient sensing to mitochondrial adaptation. Our findings provide new mechanistic insights into the metabolic regulation of the OMM proteome, which may link diet-induced changes in mitochondrial proteostasis and metabolism to diseases such as cancer and infertility.
Methods
C. elegans maintenance and transgenic lines
C. elegans were grown at 20 °C and kept on nematode growth medium (NGM) plates seeded with E. coli OP50 bacteria as food source as previously described73. The strain PP4027: unc-119(ed4)III; hhIs286[Peft-3::UbV::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ]II was generated by ballistic gene transfer of a plasmid containing a UbV::GFP::fis-1-TM fusion gene under the ubiquitous promoter eft-3. The strains PP4032: unc-119(ed4)III; hhEx203[Peft-3::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ] and PP4035 unc-119(ed4)III; hhIs287[Peft-3::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ] were generated by ballistic gene transfer of a plasmid containing a GFP::fis-1-TM fusion gene under the ubiquitous promoter eft-3. PP4028: unc-119(ed4)III; hhIs286[Peft-3::UbV::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ]II; foxSi75 [eft-3p::tomm-20::mKate2::HA::tbb-2 3′UTR]I was generated by crossing PP4027: unc-119(ed4)III; hhIs286[Peft-3::UbV::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ]II with SJZ328: foxSi75[eft-3p::tomm-20::mKate2::HA::tbb-2 3’UTR]I. PP4029: unc-119(ed4)III; foxSi75[eft-3p::tomm-20::mKate2::HA::tbb-2 3′UTR]I; hhIs64[unc-119(+); sur-5::UbV-GFP]III was generated by crossing SJZ328: foxSi75 [eft-3p::tomm-20::mKate2::HA::tbb-2 3′UTR]I with PP563: unc-119(ed4); hhIs64[unc-119(+); sur-5::UbV-GFP]III. PP4031: cdc-48.1(tm544)II; unc-119(ed4)III; hhIs286[Peft-3::UbV::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ]II was generated by crossing cdc-48.1(tm544)II with PP4027: unc-119(ed4)III; hhIs286[Peft-3::UbV::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ]II. PP4038: hhIs286[Peft-3::UbV::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ]II; risIs33 [K03A1.5p::3xFLAG::SV40-NLS::dCas9::SV40-NLS::VP64::HA + unc-119(+)] was generated by crossing MIR249: risIs33 [K03A1.5p::3xFLAG::SV40-NLS::dCas9::SV40-NLS::VP64::HA + unc-119(+)] with PP4027; PP4039: hhIs287[Peft-3::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ]; risIs33 [K03A1.5p::3xFLAG::SV40-NLS::dCas9::SV40-NLS::VP64::HA + unc-119(+)] was generated by crossing MIR249 with PP4035. PP4044: unc-119(ed3); hhIs286[Peft-3::UbV::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ]II; qaIs7201[pcdc-48.1::FLAG::cdc-48.1, unc-119(+)] was generated by crossing XA7201: unc-119(ed3); qaIs7201[pcdc-48.1::FLAG::cdc-48.1,unc-119(+)] with PP4027. The CRISPR–Cas9 strategy to generate the bcat-1(E279K) mutant strain PP4209: +/szT1[lon-2(e678) umnIs39]I;bcat-1(E279K)/szT1[umnIs40]X and the subsequent balanced strain PP4211: hhIs286[Peft-3::UbV::GFP::fis-1-TM::unc-54 3′UTR; unc-119 + ]II; +/szT1[lon-2(e678) umnIs39]I; bcat-1(E279K)/szT1[umnIs40]X were developed together with SunyBiotech and mutant isolation was performed by the company. All strains that were used in this study are listed in Supplementary Table 4.
Human cells maintenance
HEK293 cells (Sigma-Aldrich, 85120602) were maintained in DMEM (Gibco, 10566-016) and 10% FBS (VWR, 89510-186). Lung cancer cell lines were cultured in RMPI with 10% FBS. H1437 (NCI-H1437 (H1437) ATCC CRL-5872) and H1666 (NCI-H1666 (H1666) ATCC CRL-5885) were purchased directly from American Type Culture Collection (ATCC). H2030 (NCI-H2030 (H2030) ATCC CRL-5914) was a kind gift from R. Jachimowicz’s laboratory. All cells were routinely tested for mycoplasma contamination using a Mycoplasma PCR Detection kit (abmGood, G238). All cell lines tested negative for mycoplasma. For the generation of stable cell lines constitutively expressing mitoUFD, the human UbV–GFP–FIS1TM construct was cloned into the EGFP-N1 vector and transfected into HEK293 Flp-In T-REx cells (Invitrogen, R78007) using the transfection reagent FuGene HD (Promega, E2311) according to the manufacturer’s guideline. Positive clones were selected using DMEM supplemented with 10% FCS, 500 mg ml−1 penicillin–streptomycin and 500 µg ml−1 geneticin (InvivoGen, ant-gn-1). Expression of constructs was verified by western blot analysis of cell lysates.
mitoTracker staining for mitochondria
For mitochondrial staining, MitoTracker Deep Red FM (Thermo Fisher, M22426) was used. Specifically, 3.5 cm NGM plates were seeded with 100 µl OP50 2 days before the assay. A 1 mM stock solution was prepared on the day of assay and 50 µl of the 1 mM stock solution was evenly spread on top of the OP50 food lawn. The MitoTracker-treated plates were protected from light and the staining plate was left to dry for 30 min. Fifty worms on day 1 of adulthood were then transferred to the staining plate and incubated for 2 h in dark. The MitoTracker-stained worms were then transferred into M9 drops next to the food lawn of a fresh OP50-seeded plate. The worms were allowed to crawl onto the fresh OP50 food lawn and incubated for 2 h in the dark ready for imaging.
Chase assay
The chase assay in worms was performed as previously described57,74. CHX plates were prepared by spreading 50 μl of 50 mg ml−1 CHX (Sigma) in ethanol stock solution on a 3.5 cm Petri dish containing 5 ml NGM using a spatula. Ethanol was added to the plates as controls and all plates were incubated at room temperature for 30 min. Plates were then seeded with OP50 bacteria overnight at 37 °C. Then, 100 L4 stage animals (synchronized by timed egg laying) were transferred to NGM plates containing freshly prepared CHX or control plates for 0 h, 3 h, 6 h or 9 h. Worms were then collected and analysed by western blotting. For cell culture, 100 µg ml−1 CHX was used.
Western blotting
To collect worm samples, 50 animals were picked into 100 µl M9 buffer, left to settle down and then 75 µl of M9 was removed. Then 25 µl 2× SDS buffer was added, followed by boiling at 95 °C for 10 min, sonication and boiling again at 95 °C for 5 min. For each sample, 30 µl was loaded on Bis–Tris 4–12% polyacrilamide gels for electrophoresis. Proteins were transferred to Amersham Protran 0.1 NC nitrocellulose membranes (Cytiva) with a semi-dry blotting system (Bio-Rad, Trans-Blot Turbo) using NuPAGE transfer buffer (Thermo Fisher Scientific). Membranes were blocked with 5% milk (in PBST + 0.1% Tween 20) for 1 h at room temperature and incubated with the primary antibodies overnight at 4 °C with RotiBlock (Carl Roth). Secondary antibodies were incubated at room temperature for 2 h. Fluorescence detection was conducted with a Li-Cor Odyssey scanner. The following antibodies were used: GFP (Clontech, 632380, 1:5,000), tubulin (Abcam, ab52866, 1:5,000), VDAC2 (Proteintech, 11663-1-AP, 1:1,000), TOMM40 (Proteintech, 66658-1-lg, 1:2,000), MTCO1 (Abcam, ab14705, 1:2,000), SEL1L (Sigma-Aldrich, S3699, 1:1,000), SYVN1 (Cell Signalling, 14773, 1:2,000), CDC-48 (Custom antibody, Hoppe lab, 1:5,000), FLAG (Sigma-Aldrich, F7425, 1:2,500), ubiquitin (Sigma-Aldrich, 05-944, 1:3,500), SEL-1 (Custom antibody, Jarosch Lab, 1:8,000), donkey anti-rabbit (Li-Cor, 926-68073, 1:10,000), donkey anti-mouse (Li-Cor, 926-32212, 1:10,000), goat anti-mouse (Jackson ImmunoResearch Laboratories, AB2338503, 1:10,000) and goat anti-rabbit (Jackson ImmunoResearch Laboratories, AB2339150, 1:10,000).
RNAi by feeding
RNAi by feeding was performed as previously described75 with modifications. RNAi plates were supplemented with 100 μg ml−1 ampicillin and 1 mM IPTG and stored at 4 °C for no more than 1 month. Bacterial strains (RNase-deficient E. coli HT115) expressing double-stranded RNA were obtained from either Ahringer library75 or Vidal library76 and confirmed by sequencing. Bacteria were grown on LB plates with ampicillin and tetracycline at 37 °C overnight. Bacterial liquid culture with 100 μg ml−1 ampicillin was grown at 37 °C with shaking at 150 rpm for 6–8 h. The liquid culture was seeded on the plates and left to dry at room temperature for 2 days. Animals were synchronized by timed egg laying: young adults were picked onto OP50 bacteria-seeded plates to lay eggs for 4 h. After 2 days, L4 stage animals were picked onto corresponding RNAi plates and day 1 adults were imaged. This acute RNAi treatment was performed to avoid potential long-term adaptation to metabolic and mitochondrial homeostasis changes and to avoid potential developmental effects for many of the mitochondrial genes. For BioSorter quantification, L4 stage animals were washed off from OP50 plates, washed 3× by M9, and then transferred to corresponding RNAi plates. Double RNAi knockdown was performed by mixing two liquid bacterial cultures with 1:1 ratio, and the mixture of L4440 with the gene of interest was used as control.
BioSorter quantification of fluorescence
For BioSorter (Union Biometrica) quantification, synchronized day 1 adults by timed egg laying were used. The fluorescence values (GFP green) of 50–250 gated worms were collected from each independent biological replicate. Gating of day 1 adults was based on time of flight >1,000 and extinction >600. Three independent biological replicates were used for statistical analysis. A representative illustration of the gating strategy is shown in Extended Data Fig. 2b.
Fluorescence microscopy
For confocal imaging in worms, L4 stage or day 1 adult hermaphrodites were immobilized with 25 mM levamisole on a 3% agarose pad. Worms were imaged using an LSM980 Airyscan 2 equipped with Plan-Apochromat 63× /1.4 oil DIC and ZEN Connect Modul (Carl Zeiss Microscopy GmbH). For whole-worm imaging, an Axiozoom V16 (Carl Zeiss Microscopy GmbH) equipped with Axiocam 506mono and Zeiss 2.3 software (Carl Zeiss Microscopy GmbH) was used. ImageJ was used for image processing and fluorescence intensity quantification. For confocal imaging, cells were plated on glass-bottom dishes 2 days before treatment and imaging.
mitoUFD pulldown for proteomic analysis or TUBE agarose IP
Twenty thousand worms were prepared by bleaching each strain for each condition. Animals were washed three times with M9 and once with dH2O. Then, 200 µl fractionation buffer (50 mM Tris pH 7.4, 150 mM NaCl, 1× EDTA-free PI cocktail (Roche) and 1 mM Pefabloc) was added to each sample, followed by 20 strokes with a Dounce homogenizer. The worm lysate was spun down at 200g for 5 min and 800g for 10 min at 4 °C, and the supernatant was transferred to a new tube. The supernatant was centrifuged at 12,000g for 30 min at 4 °C to pellet the mitochondrial fraction. The mitochondria pellet was resuspended and solubilized in 500 µl fractionation buffer containing 1% (w/v) digitonin for 45 min at 4 °C. Insoluble material was removed by centrifugation at 16,000g for 10 min at 4 °C. For mitoUFD pulldown, 50 µl GFP-Trap Magnetic Agarose (ChromoTek) was added to the supernatant. The samples were incubated at 4 °C while rotating at 10 rpm for 1 h. The sample was transferred on a magnet stand and washed with fractionation buffer containing 0.1% (w/v) digitonin, 0.5% NP40 and 0.1% Triton-100. A mild wash was performed with fractionation buffer containing 0.1% (w/v) digitonin. The pellet was resuspended in 25 µl fractionation buffer and 25 µl 10% SDS (in PBS) was added. The samples were incubated at 95 °C for 5 min and transferred to a new tube without beads. Samples were sonicated and further processed with a modified SP3 protocol77 and analysed by the Cologne Excellence Cluster on Cellular Stress Responses in Aging-Associated Diseases (CECAD) Proteomics Facility. For immunoprecipitation (IP) with TUBE1 agarose (LifeSensors), fractionation buffer was supplemented with 10 mM NEM and 25 µl supernatant was used as a pre-IP control. The rest of the sample was incubated with TUBE agarose at 4 °C while rotating at 10 rpm overnight. Beads were collected by low-speed centrifugation (1,000g) at 4 °C for 5 min. Beads were washed with 1 ml fractionation buffer containing 0.1% (w/v) digitonin, collected by low-speed centrifugation and the supernatant aspirated, leaving a small volume cushion to avoid disturbing the beads (2×). The pellet was resuspended in 25 µl fractionation buffer for western blot analysis.
Proteasome activity assay
Proteasome activity measurement was performed as previously described78 with modification. Cells were seeded on a 6-well plates 2 days before the measurement. On the day of measurement, cells were treated with 1 mM Leu or dH2O control for 3 h. Cells were collected with ice-cold PBS and washed twice. For whole-cell samples, 200 µl proteasome lysis buffer (50 mM Tris–HCl pH 7.5, 250 mM sucrose, 5 mM MgCl2, 0.5 mM EDTA, freshly added 2 mM ATP and 1 mM DTT) was added, and the cells were lysed by passing ten times through 27 G syringe. After centrifuging at 10,000g for 10 min at 4 °C, the supernatant was collected. For mitochondria samples, cells were resuspended in 200 µl MTiso buffer (3 mM HEPES pH 7.4, 210 mM mannitol, 70 mM sucrose, 0.2 mM EGTA and freshly added 1× EDTA-free protease inhibitor) and transferred into an ice-cold Dounce homogenizer. Fifty strokes were applied and the homogenate was piled up to 200 µl of 340 mM sucrose. After centrifugation at 500g for 5 min at 4 °C, the supernatant was collected and followed by centrifugation at 10,000g for 10 min at 4 °C. The pellet was resuspended in 200 µl proteasome lysis buffer. The following AMC substrates were used: Trypsin-like proteasome activity: Ac-Arg-Leu-Arg-AMC (Enzo, BWL-AW9785-0005); Chymotrypsin-like proteasome activities: Z-Gly-Gly-Leu-AMC (Enzo, BML-ZW8505-0005); and Caspase-like proteasome activities: Z-Leu-Leu-Glu-AMC (Enzo, BWL-ZW9345-0005)
OMM protein isolation
Isolation of OMM proteins was performed as described previously with modification79,80. Cells were seeded 1 day before collection. For each sample, four 151 cm2 dishes were seeded with with 5.7 × 106 cells per dish. On the day of collection, cells were treated with the corresponding reagents for 3 h. GCN-2 inhibitor (HY-100877) was purchased from MedChemExpress. Cells were collected and washed twice with PBS. Cells were resuspended in 5 ml MTiso buffer as described before (3 mM HEPES pH 7.4, 210 mM mannitol, 70 mM sucrose, 0.2 mM EGTA and 1× EDTA-free protease inhibitor). Cells were transferred into an ice-cold Dounce homogenizer followed by 50 strokes. Then 500 µl was removed as the whole-cell control and the homogenate was piled up in a 50 ml Falcon to an equal volume of 340 mM sucrose, centrifuged at 500g at 4 °C for 5 min and the supernatant collected. The samples were then centrifuged at 4 °C for 10 min at 10,000g to pellet mitochondria. The mitochondria pellet was resuspended with 500 µl MTiso buffer containing 1 mg ml−1 digitonin and 10 mM NEM. The samples were mixed intensely in a thermo mixed at 4 °C, 400 rpm for 15 min and 500 µl MTiso buffer added to stop digitonin extraction followed by centrifugation at 4 °C, 10,000g for 10 min to collect the supernatant, which contains solubilized OMM and intermembrane space proteins. Protein concentration was measured by the BCA assay (Pierce) and the same amount of proteins were loaded for each sample for western blot analysis.
Fractionation assay
A fractionation assay was performed as previously described79. The cells were collected and washed twice with ice-cold PBS. Cells were resuspended with 500 µl MTiso buffer followed by 50 strokes with a Dounce homogenizer. The homogenate was piled up to an equal volume of 340 mM sucrose, centrifuged at 500g at 4 °C for 5 min and the supernatant collected. The samples were centrifuged at 4 °C for 10 min at 10,000g to pellet mitochondria. The supernatant was centrifuged at 15,000g for 20 min and collected, then centrifuged at 100,000g at 4 °C for 1 h to pellet the microsomes. Concentrations of mitochondria and microsomes were measured by the BCA assay.
BTZ treatment in worms
BTZ stock was diluted in DMSO at 10 mM. Then 3.5 cm plates seeded with OP50 were supplemented with dH2O-diluted BTZ or DMSO control to a final concentration of 5 µM and 10 µM. The plate was left to dry for 2 h. L4 stage animals were transferred to BTZ or control plates for 6 h and immediately used for fluorescence imaging or western blotting.
Leu supplementation
Leu stock solution was prepared at 100 mM diluted in dH2O. Synchronized L4 stage animals were washed off from OP50-seeded plates to a 1.5 ml Eppendorf tube. Heat-killed OP50 was prepared as follows: OP50 culture was prepared by shaking at 37 °C for 6 h. The culture was then concentrated 20× and diluted in M9 followed by incubation at 75 °C for 90 min. The heat-killed OP50 was cooled down to room temperature before use. In a 100 µl liquid culture, 40 µl HK-OP50 and 3–5 µl 20× concentrated live OP50 was added. Then 100 mM Leu stock solution was added to reach the final concentration of 20 mM and 50 mM. dH2O was used as a control. Worms were incubated with gentle shaking for 3 h before the assay. For the RNAi + Leu supplementation assay, animals were cultured on corresponding RNAi plates from the L2 to L4 stage, and Leu supplementation was performed on L4 stage animals. The corresponding RNAi bacteria culture was heat killed and used for the assay.
Overexpression by feeding
Overexpression of genes of interest by bacterial feeding was performed according to a previous publication81 with slight modifications. L4440_gcn-2_sgRNA were constructed using L4440_BioBrick-sgRNA as backbone. L4440_BioBrick-sgRNA was digested with BbsI (NEB) and gcn-2_gRNA_A fragments (annealed AGGGAAACAAGCGCCAAAAAGTGG and AAACCCACTTTTTGGCGCTTGTTT) were inserted using T4 ligation (NEB) (vector: insert of 1:10). L4440_gcn-2_gRNA_A was digested using BsaI HF-V2 (NEB) and the gcn-2_gRNA_B fragments (annealed AGGGAGAGGTTCCAACTAATCAAG and AAACCTTGATTAGTTGGAACCTCT) were inserted using T4 ligation (NEB). Both L4440_gcn-2_sgRNA and L4440_SCR_sgRNA (control) were transformed to HT115 bacteria for worm feeding. The feeding procedure was performed as described in ‘RNAi by feeding’. Overexpression of gcn-2 was validated by qPCR.
RNA extraction, cDNA synthesis and qPCR
Total RNA isolation was performed using TRIzol (Invitrogen). Age-synchronized worms were collected and washed twice with M9 buffer. Then 1 ml of TRIzol was then added to the worm pellet. Worms were frozen at −80 °C for overnight and then thawed at 37 °C. Zirconia beads were added for the Precellys 24-Dual cell homogenizer (Peq-Lab) disruption twice for 20 s at 6,000 rpm. Samples were incubated for 5 min at room temperature. Next, 300 µl 1-bromo-3-chloropropane was added followed by shaking for 15 s. Samples were then incubated at room temperature for 2–3 min and centrifuged for 15 min at 12,000g at 4 °C to separate the aqueous and organic phase. The aqueous phase was used to isolate total RNA with the RNeasy Mini kit (Qiagen) following the manufacturer’s instructions. The quality and concentration of the isolated RNA was measured using a NanoDrop 8000 spectrophotometer (Thermo Fisher Scientific). A total of 1,000–2,000 ng of total RNA was used for cDNA synthesis with the High-Capacity cDNA Reverse Transcription kit (Applied Biosystems) following the manufacturer’s instructions. qPCR was performed with Luna Universal qPCR Master Mix (New England Biolabs) and the Bio-Rad CFX96 Real-Time PCR Detection System. Three technical replicates were analysed per sample. gpd-1 was used as reference for normalization. The following primers were used: gcn-2 F: CAAATAGTACTTGACGAACGGGTA; gcn-2 R: CACCCAGACATGCCAATGAG; gpd-1 F: ATCACGTTGTTTCTAACGCATC; gpd-1 R: ATGAGTCCTTCGATGATACCG
Proteomic analysis of whole-worm samples and enriched mitochondria
For proteomic analysis in C. elegans, 10k worms (N2) were used for each sample. Worms were synchronized by egg preparation with bleaching. Leu (final concentration of 20 mM) and/or CHX (final concentration of 500 µg ml−1) were supplemented to L4 stage animals in HK-OP50 for 3 h with dH2O as a control treatment. Worms were then washed 3× with M9 and 1× with ddH2O. For the worm pellet, 200 ml fractionation buffer (50 mM Tris pH 7.4, and 150 mM NaCl; before use add 1× EDTA-free PI cocktail and 1 mM Pefabloc) was added followed by 20 stokes with a Dounce homogenizer. Then 25 µl was removed as whole-worm samples and the remainder of the worm lysate was spun down at 200g for 5 min and 800g for 10 min at 4 °C. The supernatant was transferred to fresh Eppendorf tubes and centrifuged at 20,000g for 60 min at 4 °C. The pellet was resuspended in 25 µl fractionation buffer and 25 µl 2× SP3 buffer (10% SDS in 1× PBS). For whole-worm samples, 25 µl of 2× SP3 buffer was added to each sample. Both whole-worm and pellet samples were then incubated at 95 °C for 5 min followed by sonification. For all samples, DTT was added to a final concentration of 5 mM, vortexed and incubated at 55 °C for 30 min. Chloroacetamide was added to a final concentration of 40 mM, vortexed and incubated in the dark for 30 min at room temperature and then centrifuged for 10 min at 20,000g. The supernatent was transferred to a new tube when a pellet was visible. Samples were further processed with a modified SP3 protocol77 and analysed by the CECAD Proteomics Facility on an Orbitrap Exploris 480 (granted by the German Research Foundation under INST 216/1163-1 FUGG) mass spectrometer coupled to a Vanquish neo in trap-and-elute set up (Thermo Scientific). The system was equipped with a FAIMSduo differential ion mobility device running at a compensation voltage of −50 V (Thermo Scientific) and an electrode temperatures of 99.5 °C (inner) and 85 °C (outer electrode). Samples were loaded onto a precolumn (Acclaim 5 µm PepMap 300 µ cartridge) with a flow of 60 µl min−1 before being reverse-flushed onto an in-house packed analytical column (30 cm length, 75 µm inner diameter, filled with 2.7 µm Poroshell EC120 C18, Agilent). Peptides were chromatographically separated with an initial flow rate of 400 nl min−1 and the following gradient: initial 2% B (0.1% formic acid in 80% acetonitrile), up to 6% in 4 min. Then, flow was reduced to 300 nl min−1 and B increased to 20% B in 50 min, up to 35% B within 27 min and up to 95% solvent B within 1.0 min while again increasing the flow to 400 nl min−1, followed by column wash with 95% solvent B and re-equilibration to initial conditions. MS1 scans were acquired from 399 m/z to 1,001 m/z at 15k resolution. Maximum injection time was set to 22 ms and the AGC target to 100%. MS2 scans ranged from 400 m/z to 1,000 m/z and were acquired at 15k resolution with a maximum injection time of 22 ms and an AGC target of 100%. DIA scans covering the precursor range from 400 to 1,000 m/z were acquired in 60 × 10 m/z windows with an overlap of 1 m/z. All scans were stored as centroid.
Proteomic analysis of whole-cell samples and enriched mitochondria
For whole-cell quantitative label-free proteomics, HEK293 Flp-In T-Rex WT cells were seeded on 6-well plates. At 2 days after seeding, the cells were treated with 1 mM l-Leu and 6.67 µl ml−1 ddH2O, respectively. For CHX-chase samples, 100 µg ml−1 CHX were added during the l-Leu/ ddH2O treatment. After incubating the cells with the respective treatments for 3 h, cells were washed with 1 ml PBS per well and collected in 0.7 ml PBS. Subsequently, cells were centrifuged for 7 min at 300g and 4 °C. The supernatant was removed and cell pellets were suspended in 50 µl lysis buffer per well (4% SDS in PBS supplemented with protease inhibitors). Cell lysates were sonicated and incubated at 96 °C for 5 min. Isolation of crude mitochondria from HEK293 cells was performed as previously described82. In short, cells were seeded on 15 cm dishes and cultivated for 3 days. Then, cells were treated with 1 mM l-Leu and 6.67 µl ml−1 ddH2O, respectively, and incubated for 3 h. For collection, the cells were washed with 10 ml ice-cold PBS per dish and collected in 10 ml ice-cold PBS. Subsequently, cells were centrifuged for 5 min at 500g and 4 °C. The supernatant was removed and cell pellets were suspended in 5 ml M buffer (220 mM mannitol, 70 mM sucrose, 5 mM HEPES–KOH and 1 mM EGTA–KOH, pH 7.4) containing cOmplete Protease Inhibitor Cocktail. Cells were homogenized using a precooled potter homogenizer (1,000 rpm, 15 strokes). The cell homogenate was centrifuged for 5 min at 600g and 4 °C. The supernatant was distributed to 2 ml reaction tubes and centrifuged again for 5 min at 600g and 4 °C. Afterwards, the supernatant containing the crude mitochondria was centrifuged for 10 min at 8,000g and 4 °C. The pellet was washed with 2 ml ice-cold M buffer (without protease inhibitor cocktail) and centrifuged 10 min at 6,000g and 4 °C. Next, the supernatant was removed, and the pellet was suspended in 400 µl ice-cold M buffer (without protease inhibitor cocktail). Then, the protein content was determined using the BCA Reagent ROTI Quant assay according to the manufacturer´s instructions. Next, 100 µg mitochondria per replicate were added to fresh low-binding reaction tubes and centrifuged for 5 min at 10,000g and 4 °C. The supernatant was removed and the pellet containing crude mitochondria was suspended in 50 µl lysis buffer (4% SDS in PBS supplemented with protease inhibitors). Crude mitochondria lysates were sonicated and incubated at 96 °C for 5 min. From this point, whole-cell and mitochondrial samples were handled in parallel and treated using the same experimental procedure. First, lysates were subjected to an acetone precipitation: 200 µl ice-cold acetone were added and samples were stored at -80 °C overnight. The following day, samples were thawed on ice and centrifuged for 15 min at 16,000g and 4 °C. The supernatant was removed and the pellet was washed with 500 µl ice-cold acetone and subsequently air dried. Afterwards, pellets were solved in 50 µl 8 M urea in TEAB buffer supplemented with protease inhibitors and sonicated. The protein concentration of the samples was determined using Pierce 660 nm protein assay reagent following the manufacturer’s instructions. Then, 50 or 30 µg (for whole-cell and mitochondrial samples, respectively) of each sample was transferred to a fresh low-binding reaction tube and the volume was adjusted to 40 µl by the addition of 8 M urea in TEAB buffer supplemented with protease inhibitors. Next, DTT was added to a final concentration of 5 mM and samples were incubated for 1 h at 25 °C. Then chloroacetamide was added to a final concentration of 40 mM and samples were incubated for 30 min in the dark at room temperature. Afterwards LysC protease was added in an enzyme to substrate ratio of 1:200 and samples were incubated for 4 h at 25 °C. Next, 160 µl of 50 mM TEAB buffer were added to reduce the urea concentration to below 2 M. After the addition of Trypsin in an enzyme to substrate ratio of 1:75, samples were incubated overnight at 25 °C. The next day, digestion was stopped by addition of formic acid to a final concentration of 1%. Finally, samples were centrifuged for 5 min at 20,000g and room temperature and loaded onto SDB-RP StageTips, which were previously equilibrated with 20 µl methanol, followed by 20 µl buffer B (0.1% formic acid in ddH2O) and two times 20 µl buffer A (0.1% formic acid in 80% acetonitrile). Samples on StageTips were washed once with 40 µl buffer A and twice with 40 µl buffer B. Afterwards, the StageTips were dried and stored at 4 °C until measurement. Samples were analysed by the CECAD Proteomics Facility on an Orbitrap Exploris 480 (granted by the German Research Foundation under INST 216/1163-1 FUGG) mass spectrometer coupled to a Vanquish neo in trap-and-elute set up (Thermo Scientific). Project-specific gas-phase fractionation libraries83 were generated by injecting individual pools stemming from the respective samples six times each covering the range of 400 to 1,000 m/z in 100 m/z steps running with an MS1 resolution of 60k and an MS2 resolution of 30k with staggered 4 m/z windows, resulting in effective 2 m/z windows after deconvolution using ProteoWizard software84.
Data processing for proteomic analysis
Samples were analysed in DIA-NN 1.8.1 (ref. 85). The WormBase database (PRJNA13758.WBPS15, downloaded 24 January 2023) was used for C. elegans data analysis. The Uniprot canonical Human database (UP5640, downloaded 09 January 2024) was used for HEK293 cell data analysis. For all analysis, DIA-NN was run with the additional command line prompts ‘—report-lib-info’. Further output settings were filtered at 0.01 false discovery rate (FDR), N-terminal methionine excision enabled, maximum number of missed cleavages set to 1, min peptide length set to 7, max peptide length set to 30, min precursor m/z set to 400, max precursor m/z set to 1,000, cysteine carbamidomethylation enabled as a fixed modification and heuristic protein inference activated. For the first analysis of Leu effect only in C. elegans, library building was performed with settings matching the acquisition parameters and the match-between-runs function enabled. Here, samples were directly used to refine the simulated FASTA-based library for a second search of the sample data. For the remaining datasets, the six gas-phase fractionation runs were first used to generate dedicated project libraries using identical settings as described above. Afterwards, samples were analysed against the corresponding library, again with identical settings as described above. Afterwards, DIA-NN output was further filtered on library q value and global q value ≤0.01 and at least two unique peptides per protein using R (4.1.3). Finally, label-free quantification values were calculated using the DIA-NN R package. Afterwards, analysis of results was performed in Perseus 1.6.15 (ref. 86). For analyses from C. elegans samples, the resulting WormBase identifiers were translated into the UniProt identifier and these were used for term annotations and further analyses. GO term analysis was performed by GOrilla87 with a ranked list of proteins according to t-test statistics. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE88 partner repository with the dataset identifiers PXD051398, PXD051401, PXD051403, PXD062734 and PXD062736.
Seahorse O2 consumption rate measurement
The oxygen consumption rate (OCR) was measured by a Seahorse XFe96 Analyser (Agilent) as previously described for worms and human cells89,90 with modifications. Worm RNAi or overexpression feeding were performed at the L2 stage (synchronized by timed egg laying) for 24 h, followed by Leu supplementation for 3 h before the assay. We used a 2 min mix, 30 s wait, 2 min measure cycle protocol for all measurements. Fifteen worms were picked into each well and bacteria-only wells were used as control. Five basal measurements were taken before the injection of the mitochondrial uncoupler carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (FCCP) (40 µM final concentration). After nine measurements, NaN3 (50 mM final concentration) was injected to inhibit mitochondrial complex IV and V activity for a further four measurements. Six technical replicates and four biological replicates were measured for each condition. Data were normalized to worm number. For HEK293 cells, 3000 cells per well were seeded 1 day before the assay and cells were treated with corresponding amino acids for 3 h before the assay. We used 1.5 µM oligomycin, 8 µM FCCP and 0.5 µM rotenone + antimycin. For measurement, we used a 3 min mix, 3 min measure cycle protocol, with three measurement cycles for each step. Data were normalized to cell number.
Immunogold labelling of frozen rehydrated C. elegans
Five adult worms were transferred into a flat carrier with a 100 µm recess (Leica, 1093), which was filled with 20% polyvinylpyrrolidone in ddH2O. The carrier was high-pressure frozen using an EMPact 2 (Leica) and stored in liquid nitrogen until freeze substitution. The samples were then incubated in a solution of 0.5% uranyl acetate, 2% ddH2O and 0.5% glutaraldehyde in acetone at −90 °C for 8 h in the AFS2 (Leica). The temperature was then raised to −30 °C within 33 h, after which the fixative was exchanged for 0.5% glutaraldehyde and 2% ddH2O in acetone, and the samples were kept for 2 h. The temperature was then raised to −20 °C within 1.5 h. Rehydration was started using a solution of 50% 0.1 M PHEM buffer and 0.5% glutaraldehyde in acetone, raising the temperature to 0 °C within 3 h. The samples were rehydrated using 75% and 90% 0.1 M PHEM in acetone with 0.5% glutaraldehyde for 15 min each at 0 °C. Then, 100% PHEM with 0.5% glutaraldehyde was added for 16 h at 0 °C. The sample was then washed with 0.1 M PHEM for 15 min at room temperature and incubated for 30 min with 0.1 M PHEM, 0.25% glycine, 0.025% azur II (Sigma-Aldrich, 861065-25 G) and 0.025% methylene blue (Sigma-Aldrich, M9140-25G). Individual worms were placed on a layer of solidified 10% gelatin and covered with another layer of 10% gelatin. Small cubes containing the worms were cut and incubated overnight or longer in 2.3 M sucrose (Sigma-Aldrich) at 4 °C. The gelatin cubes were mounted onto aluminium pins (Leica, 16701950) for cryo-ultramicrotomy, then frozen at −115 °C inside the FC7 cryo-chamber (Leica), which was mounted to the UC6 ultramicrotome (Leica). Ultrathin cryo-sections (70 nm) were cut using a diamond knife (Diatome). The sections were picked up using wire loops in an ice-cold 1:2 mixture of 2% methylcellulose (Sigma-Aldrich) and 2.3 M sucrose and transferred onto 100 mesh formvar-coated copper grids. The gelatin was then removed by incubating the grids in PBS at 40 °C for 1 h and washing them three times with PBS. Free aldehyde groups were then inactivated with a 15 min wash in 0.05 M glycine in PBS. The grids were then incubated in a drop of blocking solution for protein A-gold (Aurion), after which they were washed three times with 0.1% BSA-C (Aurion) in PBS. The primary antibody was diluted 1:50 in 0.1% BSA-c in PBS and incubated for 90 min, followed by six rinses with 0.1% BSA-c in PBS. No primary antibody controls were performed in parallel. The antigen was detected by incubating for 90 min with protein A-gold (10 nm), diluted 1:20 in 0.1% BSA-c in PBS. After 5 min of fixation with 2% glutaraldehyde, the sections were washed in PBS and H2O, then contrasted for 5 min with 0.4% uranyl acetate in 2% methylcellulose on ice. The sections were picked up with a wire loop, with the excess fluid drained by gently dragging the loop over Whatman filter paper. After air drying, the grids were removed from the loops using forceps. Electron micrographs were taken using a JEM-2100 Plus transmission electron microscope (JEOL) equipped with a OneView camera 4 K 32-bit (Gatan) and Digital Micrograph software (Gatan).
Brood size assay
Brood size assay was performed as previously described91 with modification. Individual L4 animals were transferred to each 3.5 cm NGM plates seeded with corresponding RNAi or overexpression bacteria. Animals were transferred to new plates every day until day 3 of adulthood. Hatched animals, unhatched eggs and unfertilized oocytes were counted for each plate after 2 days of animal transfer.
Cell viability and BCAA-Glo assay
Cell viability assay was performed as previously described92. Three thousand cells were seeded per well in a 96-well plate in 200 ml medium on day 0. For H1666, 6,000 cells were seeded on day 0. For each condition each day, three wells were seeded as technical replicates. The DMSM (for HEK293) or RPMI (for lung cancer cells) medium contain the corresponding drugs indicated in each figure legends. Next, 40 µl Cell-Titer-Glo reagent (Promega) was added to each well and mixed briefly for 2 min. The plate was incubated at room temperature for 10 min and luminescence was measured with a plate reader at day 0 (5 h after plating), day 2 and day 4. For the BCAA measurement, the BCAA-Glo kit (Promega) was used. Cells were washed with cold PBS and resuspended at the concentration of 5 × 105 cells ml−1. Then 500 µl cells were lysed with 250 µl 0.6 N HCl, mixed well and incubated at room temperature for 5 min. Next, 250 µl neutralization buffer was added and 50 µl cell lysate was transferred to each well, followed by adding 50 µl freshly prepared BCAA detection reagent then mixed by shaking the plate for 1 min and incubated at room temperature for 1 h before measuring.
Statistics and reproducibility
For ImageJ quantification of fluorescence intensity, at least three independent biological replicates were performed with six to eight animals in each replicate. For BioSorter quantification of fluorescence intensity, three independent biological replicates were performed with 50–250 animals in each replicate. For statistical analysis, the percentage change was calculated as (GFP intensity of experimental animals − GFP intensity of control animals)/GFP intensity of control animals. The dashed line at 10% was used to evaluate the changes of substrate levels, in addition to the P value. Prism 10 was used for statistical analysis. Data distribution was assumed to be normal but this was not formally tested. For multiple comparison, one-way analysis of variance (ANOVA) or two-way ANOVA were performed, followed by post hoc test indicated under individual figures. For two-sample comparisons, an unpaired two-tailed t-test or Mann–Whitney U test was performed as indicated under the corresponding figures. Pathway analysis of amino acid genes was performed by WormFlux Pathway Enrichment Analysis (http://wormflux.umassmed.edu/WormPaths/pea.php)
All representative images from fluorescence microscopy and western blot were performed three times independently. No statistical methods were used to pre-determine sample size. No data were excluded from the analyses. The experiments were not randomized. Data collection and analysis were not performed blind to the conditions of the experiments.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifiers PXD051398, PXD051401, PXD051403, PXD062734 and PXD062736. Plasmids and C. elegans strains generated in this study will be distributed to researchers upon request. Source data are provided with this paper.
References
Metzger, M. B., Scales, J. L., Dunklebarger, M. F., Loncarek, J. & Weissman, A. M. A protein quality control pathway at the mitochondrial outer membrane. eLife 9, e51065 (2020).
Heo, J.-M. et al. A stress-responsive system for mitochondrial protein degradation. Mol. Cell 40, 465–480 (2010).
Wu, X., Li, L. & Jiang, H. Doa1 targets ubiquitinated substrates for mitochondria-associated degradation. J. Cell Biol. 213, 49–63 (2016).
Taylor, E. B. & Rutter, J. Mitochondrial quality control by the ubiquitin–proteasome system. Biochem. Soc. Trans. 39, 1509–1513 (2011).
Ravanelli, S., den Brave, F. & Hoppe, T. Mitochondrial quality control governed by ubiquitin. Front. Cell Dev. Biol. 8, 270 (2020).
Rödl, S. & Herrmann, J. M. The role of the proteasome in mitochondrial protein quality control. IUBMB Life https://doi.org/10.1002/iub.2734 (2023).
Mårtensson, C. U. et al. Mitochondrial protein translocation-associated degradation. Nature 569, 679–683 (2019).
König, T. et al. MIROs and DRP1 drive mitochondrial-derived vesicle biogenesis and promote quality control. Nat. Cell Biol. 23, 1271–1286 (2021).
Neuspiel, M. et al. Cargo-selected transport from the mitochondria to peroxisomes is mediated by vesicular carriers. Curr. Biol. 18, 102–108 (2008).
Soubannier, V. et al. A vesicular transport pathway shuttles cargo from mitochondria to lysosomes. Curr. Biol. 22, 135–141 (2012).
Hughes, A. L., Hughes, C. E., Henderson, K. A., Yazvenko, N. & Gottschling, D. E. Selective sorting and destruction of mitochondrial membrane proteins in aged yeast. eLife 5, e13943 (2016).
Schuler, M.-H. et al. Mitochondrial-derived compartments facilitate cellular adaptation to amino acid stress. Mol. Cell 81, 3786–3802.e13 (2021).
Rodriguez-Enriquez, S., Kim, I., Currin, R. T. & Lemasters, J. J. Tracker dyes to probe mitochondrial autophagy (mitophagy) in rat hepatocytes. Autophagy 2, 39–46 (2006).
Kim, I., Rodriguez-Enriquez, S. & Lemasters, J. J. Selective degradation of mitochondria by mitophagy. Arch. Biochem. Biophys. 462, 245–253 (2007).
Morgenstern, M. et al. Definition of a high-confidence mitochondrial proteome at quantitative scale. Cell Rep. 19, 2836–2852 (2017).
Neupert, W. & Herrmann, J. M. Translocation of proteins into mitochondria. Annu. Rev. Biochem. 76, 723–749 (2007).
Schmidt, O., Pfanner, N. & Meisinger, C. Mitochondrial protein import: from proteomics to functional mechanisms. Nat. Rev. Mol. Cell Biol. 11, 655–667 (2010).
Krüger, V. et al. Identification of new channels by systematic analysis of the mitochondrial outer membrane. J. Cell Biol. 216, 3485–3495 (2017).
Palmieri, F. & Pierri, C. L. Mitochondrial metabolite transport. Essays Biochem. 47, 37–52 (2010).
Colombini, M. VDAC structure, selectivity, and dynamics. Biochim. Biophys. Acta 1818, 1457–1465 (2012).
Youle, R. J. & van der Bliek, A. M. Mitochondrial fission, fusion, and stress. Science 337, 1062–1065 (2012).
Zhang, Y. et al. Synergistic mechanism between the endoplasmic reticulum and mitochondria and their crosstalk with other organelles. Cell Death Discov. 9, 51 (2023).
Song, J., Herrmann, J. M. & Becker, T. Quality control of the mitochondrial proteome. Nat. Rev. Mol. Cell Biol. 22, 54–70 (2021).
Cohen, M. M. et al. Sequential requirements for the GTPase domain of the mitofusin Fzo1 and the ubiquitin ligase SCFMdm30 in mitochondrial outer membrane fusion. J. Cell Sci. 124, 1403–1410 (2011).
Leboucher, G. P. et al. Stress-induced phosphorylation and proteasomal degradation of mitofusin 2 facilitates mitochondrial fragmentation and apoptosis. Mol. Cell 47, 547–557 (2012).
Cohen, M. M. J., Leboucher, G. P., Livnat-Levanon, N., Glickman, M. H. & Weissman, A. M. Ubiquitin–proteasome-dependent degradation of a mitofusin, a critical regulator of mitochondrial fusion. Mol. Biol. Cell 19, 2457–2464 (2008).
Belgareh-Touzé, N., Cavellini, L. & Cohen, M. M. Ubiquitination of ERMES components by the E3 ligase Rsp5 is involved in mitophagy. Autophagy 13, 114–132 (2017).
Fritz, S., Weinbach, N. & Westermann, B. Mdm30 is an F-box protein required for maintenance of fusion-competent mitochondria in yeast. Mol. Biol. Cell 14, 2303–2313 (2003).
Ota, K., Kito, K., Okada, S. & Ito, T. A proteomic screen reveals the mitochondrial outer membrane protein Mdm34p as an essential target of the F-box protein Mdm30p. Genes Cells 13, 1075–1085 (2008).
Chen, Z. et al. Mitochondrial E3 ligase MARCH5 regulates FUNDC1 to fine-tune hypoxic mitophagy. EMBO Rep. 18, 495–509 (2017).
Cao, Y. et al. A mitochondrial SCF-FBXL4 ubiquitin E3 ligase complex degrades BNIP3 and NIX to restrain mitophagy and prevent mitochondrial disease. EMBO J. 42, e113033 (2023).
Johnson, E. S., Ma, P. C. M., Ota, I. M. & Varshavsky, A. A proteolytic pathway that recognizes ubiquitin as a degradation signal. J. Biol. Chem. 270, 17442–17456 (1995).
Segref, A., Torres, S. & Hoppe, T. A screenable in vivo assay to study proteostasis networks in Caenorhabditis elegans. Genetics 187, 1235–1240 (2011).
Ahier, A. et al. Affinity purification of cell-specific mitochondria from whole animals resolves patterns of genetic mosaicism. Nat. Cell Biol. 20, 352–360 (2018).
Barthelme, D. & Sauer, R. T. Origin and functional evolution of the Cdc48/p97/VCP AAA+ protein unfolding and remodeling machine. J. Mol. Biol. 428, 1861–1869 (2016).
van den Boom, J. & Meyer, H. VCP/p97-mediated unfolding as a principle in protein homeostasis and signaling. Mol. Cell 69, 182–194 (2018).
Rath, S. et al. MitoCarta3.0: an updated mitochondrial proteome now with sub-organelle localization and pathway annotations. Nucleic Acids Res. 49, gkaa1011 (2020).
Palmieri, F. The mitochondrial transporter family SLC25: identification, properties and physiopathology. Mol. Aspects Med. 34, 465–484 (2013).
Ryan, D. G. et al. Disruption of the TCA cycle reveals an ATF4-dependent integration of redox and amino acid metabolism. eLife 10, e72593 (2021).
Owen, O. E., Kalhan, S. C. & Hanson, R. W. The key role of anaplerosis and cataplerosis for citric acid cycle function. J. Biol. Chem. 277, 30409–30412 (2002).
Martínez-Reyes, I. & Chandel, N. S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 11, 102 (2020).
Cardaci, S. et al. Pyruvate carboxylation enables growth of SDH-deficient cells by supporting aspartate biosynthesis. Nat. Cell Biol. 17, 1317–1326 (2015).
Arnold, P. K. & Finley, L. W. S. Regulation and function of the mammalian tricarboxylic acid cycle. J. Biol. Chem. 299, 102838 (2023).
Walker, M. D. et al. WormPaths: Caenorhabditis elegans metabolic pathway annotation and visualization. Genetics 219, iyab089 (2021).
Kim, J.-H. & Johnston, M. Two glucose-sensing pathways converge on Rgt1 to regulate expression of glucose transporter genes in Saccharomyces cerevisiae. J. Biol. Chem. 281, 26144–26149 (2006).
Kraakman, L. et al. A Saccharomyces cerevisiae G-protein coupled receptor, Gpr1, is specifically required for glucose activation of the cAMP pathway during the transition to growth on glucose. Mol. Microbiol. 32, 1002–1012 (1999).
Garcia-Barrio, M., Dong, J., Ufano, S. & Hinnebusch, A. G. Association of GCN1–GCN20 regulatory complex with the N-terminus of eIF2α kinase GCN2 is required for GCN2 activation. EMBO J. 19, 1887–1899 (2000).
Dong, J., Qiu, H., Garcia-Barrio, M., Anderson, J. & Hinnebusch, A. G. Uncharged tRNA Activates GCN2 by displacing the protein kinase moiety from a bipartite tRNA-binding domain. Mol. Cell 6, 269–279 (2000).
Qiu, H., Dong, J., Hu, C., Francklyn, C. S. & Hinnebusch, A. G. The tRNA-binding moiety in GCN2 contains a dimerization domain that interacts with the kinase domain and is required for tRNA binding and kinase activation. EMBO J. 20, 1425–1438 (2001).
Brüggenthies, J. B. et al. A cell-based chemical-genetic screen for amino acid stress response inhibitors reveals torins reverse stress kinase GCN2 signaling. J. Biol. Chem. 298, 102629 (2022).
Dodd, K. M. & Tee, A. R. Leucine and mTORC1: a complex relationship. Am. J. Physiol. Endocrinol. Metab. 302, E1329–E1342 (2012).
Ravanelli, S. et al. Reprograming of proteasomal degradation by branched chain amino acid metabolism. Aging Cell https://doi.org/10.1111/acel.13725 (2022).
Lesnik, C. et al. Enhanced branched-chain amino acid metabolism improves age-related reproduction in C. elegans. Nat. Metab. https://doi.org/10.1038/s42255-024-00996-y (2024).
Stonyte, V. et al. The GCN2/eIF2αK stress kinase regulates PP1 to ensure mitotic fidelity. EMBO Rep. 24, e56100 (2023).
Qi, L., Tsai, B. & Arvan, P. New insights into the physiological role of endoplasmic reticulum-associated degradation. Trends Cell Biol. 27, 430–440 (2017).
Sasagawa, Y., Yamanaka, K. & Ogura, T. ER E3 ubiquitin ligase HRD-1 and its specific partner chaperone BiP play important roles in ERAD and developmental growth in Caenorhabditis elegans. Genes Cells 12, 1063–1073 (2007).
Efstathiou, S. et al. ER-associated RNA silencing promotes ER quality control. Nat. Cell Biol. 24, 1714–1725 (2022).
Chakrabarti, S., Dube, D. K. & Roy, S. C. Effects of emetine and cycloheximide on mitochondrial protein synthesis in different systems. Biochem. J. 128, 461–462 (1972).
Fujiki, M. & Verner, K. Coupling of cytosolic protein synthesis and mitochondrial protein import in yeast. Evidence for cotranslational import in vivo. J. Biol. Chem. 268, 1914–1920 (1993).
Sasagawa, Y. et al. Caenorhabditis elegans p97 controls germline-specific sex determination by controlling the TRA-1 level in a CUL-2-dependent manner. J. Cell Sci. 122, 3663–3672 (2009).
Miyata, N. et al. Pharmacologic rescue of an enzyme-trafficking defect in primary hyperoxaluria 1. Proc. Natl Acad. Sci. USA 111, 14406–14411 (2014).
Lionaki, E. et al. Mitochondrial protein import determines lifespan through metabolic reprogramming and de novo serine biosynthesis. Nat. Commun. 13, 651 (2022).
Wang, X. L., Li, C. J., Xing, Y., Yang, Y. H. & Jia, J. P. Hypervalinemia and hyperleucine-isoleucinemia caused by mutations in the branched-chain-amino-acid aminotransferase gene. J. Inherit. Metab. Dis. 38, 855–861 (2015).
Xu, E., Ji, B., Jin, K. & Chen, Y. Branched-chain amino acids catabolism and cancer progression: focus on therapeutic interventions. Front. Oncol. 13, 1220638 (2023).
Kalita, B. et al. PAX translocations remodel mitochondrial metabolism through altered leucine usage in rhabdomyosarcoma. Cell https://doi.org/10.1016/j.cell.2025.03.008 (2025).
Abdullah, M. O. et al. Mitochondrial hyperfusion via metabolic sensing of regulatory amino acids. Cell Rep. 40, 111198 (2022).
Liang, C., Curry, B. J., Brown, P. L. & Zemel, M. B. Leucine modulates mitochondrial biogenesis and SIRT1-AMPK signaling in C2C12 myotubes. J. Nutr. Metab. 2014, 239750 (2014).
Wei, X. et al. Proteomic screens of SEL1L-HRD1 ER-associated degradation substrates reveal its role in glycosylphosphatidylinositol-anchored protein biogenesis. Nat. Commun. 15, 659 (2024).
Zhou, Z. et al. Endoplasmic reticulum-associated degradation regulates mitochondrial dynamics in brown adipocytes. Science 368, 54–60 (2020).
Liao, P.-C. & Pon, L. A. Analysis of the mitochondria-associated degradation pathway (MAD) in yeast cells. Methods Enzymol. 707, 585–610 (2024).
Stolz, A., Hilt, W., Buchberger, A. & Wolf, D. H. Cdc48: a power machine in protein degradation. Trends Biochem. Sci. 36, 515–523 (2011).
Karbowski, M. & Youle, R. J. Regulating mitochondrial outer membrane proteins by ubiquitination and proteasomal degradation. Curr. Opin. Cell Biol. 23, 476–482 (2011).
Brenner, S. The genetics of Caenorhabditis elegans. Genetics 77, 71–94 (1974).
Segref, A. et al. Thermosensation in Caenorhabditis elegans is linked to ubiquitin-dependent protein turnover via insulin and calcineurin signalling. Nat. Commun. 13, 5874 (2022).
Kamath, R. S. et al. Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature 421, 231–237 (2003).
Rual, J.-F. et al. Toward improving Caenorhabditis elegans phenome mapping with an ORFeome-based RNAi library. Genome Res. 14, 2162–2168 (2004).
Hughes, C. S. et al. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 14, 68–85 (2019).
Vilchez, D. et al. RPN-6 determines C. elegans longevity under proteotoxic stress conditions. Nature 489, 263–268 (2012).
Pallotti, F. & Lenaz, G. Isolation and subfractionation of mitochondria from animal cells and tissue culture lines. Methods Cell. Biol. 80, 3–44 (2007).
Nishimura, N., Gotoh, T., Oike, Y. & Yano, M. TMEM65 is a mitochondrial inner-membrane protein. PeerJ 2, e349 (2014).
Fischer, F. et al. Ingestion of single guide RNAs induces gene overexpression and extends lifespan in Caenorhabditis elegans via CRISPR activation. J. Biol. Chem. 298, 102085 (2022).
Murschall, L., Peker, E., MacVicar, T., Langer, T. & Riemer, J. Protein import assay into mitochondria isolated from human cells. Bio Protoc. 11, e4057 (2021).
Searle, B. C. et al. Generating high quality libraries for DIA MS with empirically corrected peptide predictions. Nat. Commun. 11, 1548 (2020).
Chambers, M. C. et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 30, 918–920 (2012).
Demichev, V., Messner, C. B., Vernardis, S. I., Lilley, K. S. & Ralser, M. DIA-NN: neural networks and interference correction enable deep proteome coverage in high throughput. Nat. Methods 17, 41–44 (2020).
Tyanova, S. et al. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 13, 731–740 (2016).
Eden, E., Navon, R., Steinfeld, I., Lipson, D. & Yakhini, Z. GOrilla: a tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinform. 10, 48 (2009).
Perez-Riverol, Y. et al. The PRIDE database resources in 2022: a hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 50, D543–D552 (2021).
Koopman, M. et al. A screening-based platform for the assessment of cellular respiration in Caenorhabditis elegans. Nat. Protoc. 11, 1798–1816 (2016).
Gu, X., Ma, Y., Liu, Y. & Wan, Q. Measurement of mitochondrial respiration in adherent cells by Seahorse XF96 cell mito stress test. STAR Protoc. 2, 100245 (2021).
Kwah, J. K. & Jaramillo-Lambert, A. Measuring embryonic viability and brood size in Caenorhabditis elegans. J. Vis. Exp. https://doi.org/10.3791/65064 (2023).
Birsoy, K. et al. An essential role of the mitochondrial electron transport chain in cell proliferation is to enable aspartate synthesis. Cell 162, 540–551 (2015).
Acknowledgements
We thank the Caenorhabditis Genetics Center (funded by NIH Office of Research Infrastructure Programs (no. P40 OD010440) and the National BioResource Project) for worm strains. We thank the CECAD imaging facility for confocal imaging (C. Jüngst) and immunogold-EM analysis (F. Gaedke and B. Martiny) (funding for instrumentation JEOL JEM-2100 Plus DFG-INST 216793-1 FUGG) and CECAD proteomics facility (supported by the large instrument grant INST 216/1163-1 FUGG by the German Research Foundation (DFG Großgeräteantrag) for the proteomics analysis. We thank G. Stellbrink and A. Seiler for technical support, S. Efstathiou for advice on BioSorter fluorescence quantification, L. Müller for support on cellular fractionation experiments, A. Segref for suggestion on CHX chase assay, R. Jachimowicz for providing and suggestions on lung cancer cell lines, M. Krüger for suggestions on proteomic data analysis and A. Correns and S. Balmert for the preliminary test of the OMM TM domain. We thank A. Andersen, N. Aravindan, S. Efstathiou, A. Franz, C. Frezza, E. Jarosch, F. Ottens, N. Podvalnaya and M. Rapé for critical comments and helpful suggestions on the paper. We also thank all members of the Hoppe lab for helpful suggestions throughout this project. This work was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) under Germany’s Excellence Strategy (EXC 2030–390661388, SFB 1218, project no. 269925409; SFB 1678, project no. 520471345; SPP 2453, project no. 541742459; RU 5762, project no. 531902955) and by the European Research Council (ERC, CellularPQCD, no. 101141579) to T.H. Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union or the European Research Council. Neither the European Union nor the granting authority can be held responsible for them. Q.L. is supported by a postdoctoral fellowship from the Alexander von Humboldt Foundation.
Funding
Open access funding provided by Universität zu Köln.
Author information
Authors and Affiliations
Contributions
Q.L. designed, performed and analysed the experiments. K.W. performed proteomic experiment and analysis in HEK293 cells and generated the human mitoUFD cell line. F.N. contributed to the establishment of CRISPR-based gene overexpression by feeding experiment. J.R. supervised the HEK293 proteomics and generation of the human mitoUFD cell line. T.H. supervised the experimental design and data interpretation. Q.L. and T.H. wrote the paper. All authors discussed the results and commented on the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Cell Biology thanks Mariusz Karbowski and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Inhibition of UPS and CDC-48 complex impairs mitoUFD degradation.
a-b. Whole worm images showing that inhibition of proteasome by bortezomib (btz) treatment increases GFP level in worms expressing mitoUFD (a) but not mitoGFP (b). c. Whole worm images showing that rpn-6.1 OE enhances mitoUFD degradation. d. ImageJ quantification showing that rpn-6.1 OE enhances the degradation of mitoUFD. Mean ± SEM, n = 3 independent biological replicates with a of 18-24 animals. ImageJ quantification. Unpaired two-tailed t-test. e. Model of Cdc48 dependent OMM protein degradation by the 26S proteasome as described in yeast. f. Western blot analysis showing that RNAi knockdown of cdc-48 as well as its cofactors ufd-3, npl-4 or ufd-1 stabilizes mitoUFD but not mitoGFP. g. Confocal images showing that RNAi knockdown of cdc-48 as well as its cofactors ufd-3, npl-4, or ufd-1 stabilizes mitoUFD at OMM. Scale bar = 10 µm. h. Worm sorter quantification showing that RNAi knockdown of cdc-48 as well as its cofactors ufd-3, npl-4, or ufd-1 increases mitoUFD levels. Mean ± SEM, n = 3 independent biological replicates with 50-250 animals in each replicate. One-way ANOVA with Holm-Sidak correction for multiple comparisons with L4440 control. Source numerical data and unprocessed blots are available in Source data.
Extended Data Fig. 2 TCA cycle affects OMM proteostasis.
a. Experimental procedure of BioSorter quantification of GFP levels in worms following RNAi knockdown of genes of interest. b. Representative profile of BioSorter quantification of whole worm fluorescence intensity. The red dots were gated for day 1 adults for fluorescence quantification based on TOF > 1000 and Extinction > 600. c. Changes in mitoUFD and cytoUFD levels after RNAi knockdown of mitochondrial genes quantified by BioSorter. n = 3 independent biological replicates with 50-250 animals in each replicate. d. TCA cycle and the C. elegans orthologues of key enzymes. e. RNAi knockdown of enzymes in TCA cycle in worms expressing mitoUFD. Mean ± sem, n = 4 independent biological replicates with total animal number of 24-32. ImageJ quantification. One way ANOVA with Holm-Sidak correction for multiple comparisons. Source numerical data are available in Source data.
Extended Data Fig. 3 RNAi screen for genes involved in amino acid metabolism, amino acid transport, glucose sensing and metabolite transport.
a. RNAi screen with the mitoUFD substrate for genes encoding enzymes for amino acid metabolism. Fluorescence in worms was quantified by worm sorter. 50-250 animals were measured for each gene. b. RNAi knockdown in the mitoUFD strain for genes involved in amino acid transportation, glucose sensing, and metabolite transportation at IMM. Mean ± sem, n = 4 independent biological replicates with total animal number of 24-32. ImageJ quantification. One way ANOVA with Dunnett correction for multiple comparisons. c. For the genes that stabilized mitoUFD of more than 20% in the targeted RNAi screen for 135 amino acid metabolic genes, the involved amino acids, ratio (hits/total genes per category) and their stabilization scores are shown based on pathway analysis. d. RNAi knockdown in the mitoGFP strain for genes involved in leucine sensing. Mean ± sem, n = 4 independent biological replicates with total animal number of 24-32. ImageJ quantification. One-way ANOVA with Dunnett correction for multiple comparisons with L4440 RNAi control. Source numerical data are available in Source data.
Extended Data Fig. 4 Leucine affects mitoUFD degradation.
a. Confocal imaging showing mitoUFD stabilisation upon RNAi knockdown of gcn-2 and bcat-1. Scale bar = 10 µm. b. Confocal imaging showing mitoUFD stabilization after 3 h leucine treatment at 20 mM and 50 mM. Scale bar = 10 µm. c. Confocal imaging showing mitoUFD expression in HEK293 cells. mitoUFD in green and mitoTracker in red. Scale = 10 µm. d. Confocal imaging showing that bortezomib treatment at 0.2 µM and 1 µM stabilise mitoUFD.
Extended Data Fig. 5 Proteasomal activity is not affected by leucine.
a-b. Activity of cytosolic proteasome or proteasome associated with mitochondria. Mean ± sem, n = 3 independent biological replicates, unpaired two-tailed t-test. c. Experimental procedure of OMM protein isolation. Mitochondria were enriched followed by OMM protein isolation. The isolated OMM proteins were confirmed by the presence of the OMM protein VDAC2 and the absence of IMM protein MTCO1. Source numerical data and unprocessed blots are available in Source data.
Extended Data Fig. 6 Additional evidence supporting the role of SEL-1 in OMM protein degradation.
a. Volcano plots showing proteomic analysis of leucine effect on whole worm. Different from enriched mitochondria, SEL-1 protein level was not affected by leucine treatment in whole worm samples. P values were determined by unpaired two-tailed t-test. b. SEL-1 and SEL-11 mediated the degradation of ERAD substrate CPL-1*. Mean ± sem, n = 3 independent biological replicates with total animal number of 18-24. ImageJ quantification. c. Leucine supplementation did not alter the levels of CPL-1*. Mean ± sem, n = 4 independent biological replicates with total animal number of 24-32. ImageJ quantification. Unpaired two-tailed t test. ns, p > 0.05. d. Fractionation assay in worms showed that SEL-1 is membrane bound and co-fractionate with MTCO1 and mitoUFD. e. Pulldown of mitoUFD and mitoGFP (control) showed that SEL-1 and SEL-11 bind to mitoUFD and mitoGFP, but the amount of SEL-1 and SEL-11 that binds to mitoUFD is more than two-fold compared to mitoGFP. f. Western blot analysis of mitoUFD substrate with enriched mitochondrial from worms treated with L4440, cdc-48, sel-1 and sel-11 RNAi. g-i. Immunogold analysis showing OMM (g) and ER (h) localization of SEL-1 in worms. No specific staining was observed in control sections lacking primary antibody (i). j. Quantification of gold particles on the mitochondrial surface in SEL-1 antibody-treated and no-antibody control samples. Source numerical data and unprocessed blots are available in Source data.
Extended Data Fig. 7 Leucine effect on mitochondrial proteome.
a. Percentage of proteins increased or decreased in whole worm proteome, MitoCarta 3.0 proteins, and OMM proteins upon leucine treatment. b-c. Volcano plots showing changes of OMM proteins (b) and whole worm and MitoCarta 3.0 proteins (c) upon leucine treatment. P values were determined by unpaired two-tailed t-test. OMM (b) and mitochondrial (c) proteins show the tendency to be more abundant in leucine-treated worms d. Log2 fold change (FC) of whole worm proteome, MitoCarta 3.0 proteins, and OMM proteins comparing Leu with dH2O control treatment. Two-tailed Mann-Whitney U test between whole worm proteins and other protein groups. e. Enriched GO terms (p < 0.001 and FDR < 0.2 cutoff) for upregulated proteins of worm mito proteome upon leucine treatment. f-g. Volcano plot showing changes of OMM proteins (f) and whole worm and MitoCarta 3.0 proteins (g) upon leucine treatment. h. Volcano plot showing changes of whole worm and MitoCarta 3.0 proteins upon CHX treatment. i-j. Volcano plot showing changes of OMM proteins (i) and whole cell and MitoCarta 3.0 proteins (j) upon leucine treatment. k. Volcano plot showing changes of whole cell and MitoCarta 3.0 proteins upon CHX treatment. P values were determined by unpaired two-tailed t-test (f-k). Source numerical data are available in Source data.
Extended Data Fig. 8 Validation of the OMM candidate TOMM40 from the proteomics.
a. Western blot for the OMM protein TOMM40 in HEK293 cells with and without 1 mM leucine treatment for 3 h. Mean ± sem, n = 12 independent biological replicates, unpaired two-tailed t test. b. CHX chase assay for the OMM protein TOMM40. The inner mitochondria membrane localised MTCO1 as control. Cells treated with 0 mM and 1 mM leucine were compared. Mean ± sem, n = 4 independent biological replicates, Two-way ANOVA, p value indicates leucine treatment effect. Source numerical data and unprocessed blots are available in Source data.
Extended Data Fig. 9 Leucine enhances mitochondrial respiration via ubiquitin-dependent remodeling of OMM proteome by SEL-1-SEL-11 and mitochondrial protein import.
a. Average O2 consumption rate by Seahorse analysis in HEK293 cells treated with essential amino acids at 1 mM for 3 hours. 3000 cells were seeded the day before assay. Mean ± sem, n = 3 independent biological replicates. b. Statistical analysis of basal respiration, ATP linked respiration and maximal respiration rates from (a). Two-way ANOVA with Fisher’s LSD test for multiple comparisons. c-d. Validation of cdc-48.1 (c) and gcn-2 (d) overexpression by western blot and/or qPCR. Mean ± sem, unpaired two-tailed t-test. e-f. Statistical analysis for Fig. 5h, i. g-h. Seahorse analysis in HEK293 cells treated with leucine or MB-12. 3000 cells were seeded the day before assay. Mean ± sem, n = 4 independent biological replicates, two-way ANOVA with Fisher’s LSD test for multiple comparisons. i. Statistical analysis for Fig. 5j. Source numerical data and unprocessed blots are available in Source data.
Extended Data Fig. 10 Additional evidence supporting the link between BCAA catabolism and OMM proteostasis in fertility and cell proliferation under OMM perturbation.
a. Design of bcat-1(E279K) CRISPR editing strain. b. Whole worm imaging of mitoUFD in WT and bcat-1(E279K)/+ worms. Scale bar = 10 µm. c. Egg hatch rate for Fig. 5l. n = 15 (WT_L4440), 14 (WT_gcn-2 RNAi), 15 (bcat-1/ + _L4440), 14 (bcat-1/+_gcn-2 RNAi) animals. d. Egg hatch rate for Fig. 5m. n = 15 (SCR control), 16 (gcn-2 OE) animals. e. Information of lung cancer cell lines from DepMap. f. Cell viability of HEK293 over four days with 0 µM, 4 µM and 10 µM MitoBlock-12 (MB-12) treatment. Two-way repeated ANOVA. Mean ± sem, n = 3 independent biological replicates with 3 technical replicates each day each condition. g-j. Cell images of HEK293 (g), H2030 (h), H1437 (i) and H1666 (j) at day 2 and day 4 after seeding. 3000 cells seeded on day 0 for HEK293, H2030 and H1437, 6000 cells seeded on day 0 for H1666. Source numerical data are available in Source data.
Supplementary information
Source data
Source Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Fig. 1 (download PDF )
Uncropped blots.
Source Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 2 (download PDF )
Uncropped blots.
Source Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Fig. 3 (download PDF )
Uncropped blots.
Source Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download PDF )
Uncropped blots.
Source Data Extended Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 1 (download PDF )
Uncropped blots.
Source Data Extended Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 5 (download PDF )
Uncropped blots.
Source Data Extended Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 6 (download PDF )
Uncropped blots.
Source Data Extended Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 8 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 8 (download PDF )
Uncropped blots.
Source Data Extended Data Fig. 9 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 9 (download PDF )
Uncropped blots.
Source Data Extended Data Fig. 10 (download XLSX )
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, Q., Weiss, K., Niwa, F. et al. Leucine inhibits degradation of outer mitochondrial membrane proteins to adapt mitochondrial respiration. Nat Cell Biol 27, 1889–1901 (2025). https://doi.org/10.1038/s41556-025-01799-3
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41556-025-01799-3
